Interested Article - Электрический синапс

- 2021-03-15
- 2
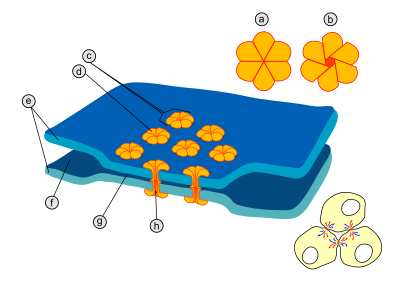
Электри́ческий си́напс ( англ. electrical synapse) — место высокоспециализированных контактов ( щелевых контактов ) между нейронами , где происходит прямое перетекание электрических токов от одного нейрона к другому . В щелевых контактах мембраны соседних клеток находятся на расстоянии около 3,8 нм , в то время как в химическом синапсе расстояние между двумя нейронами составляет от 20 до 40 нм . У многих животных в нервной системе имеются как химические, так и электрические синапсы . По сравнению с химическими синапсами, электрические синапсы проводят нервные импульсы быстрее, однако, в отличие от химических синапсов, сигнал на постсинаптическом нейроне оказывается равным или меньше первоначального сигнала. Электрические синапсы активно используют животные, которым необходима способность развивать наиболее быстрый ответ, какой возможно, например, в случае защитных рефлексов . Как правило, электрические синапсы двунаправленны, то есть нервный импульс может проходить по ним в обоих направлениях .
Структура
В области каждого щелевого контакта имеется множество особых каналов, пересекающих мембраны обеих клеток . Диаметр таких каналов составляет от 1,2 до 2 нм , поэтому через них могут проходить из одной клетки в другую ионы и молекулы среднего размера, благодаря чему цитоплазмы двух соседних клеток оказываются соединёнными . Поэтому когда мембранный потенциал одной из клеток меняется, из неё ионы могут переместиться в соседнюю клетку, деполяризуя её за счёт своего положительного заряда . Каналы щелевых контактов состоят из двух полуканалов, которые у позвоночных называются коннексонами (каждый полуканал принадлежит одной из клеток, соединённых синапсом) . Каждый коннексон образован шестью белковыми субъединицами — . Коннексин имеет длину 7,5 нм и содержит четыре участка. Коннексины в составе коннексона одинаковы или могут немного отличаться . Частным случаем электрического синапса является ( англ. autapse), в образовании которого принимают участие аксон и дендрит одного и того же нейрона .
Функции

Простота устройства электрических синапсов позволяет им проводить сигнал очень быстро, однако они участвуют лишь в простых поведенческих реакциях, в отличие от более сложно устроенных химических синапсов. Поскольку для передачи сигнала через электрический синапс не нужно связывание рецептора с сигнальной молекулой- лигандом , при работе электрических синапсов не происходит задержки, которая у химических синапсов может составлять от 0,5 до 4 миллисекунд. Однако у млекопитающих различия в скоростях проведения сигнала электрическим и химическим синапсом не различаются так сильно, как у холоднокровных животных . Благодаря высокой скорости проведения сигнала электрическим синапсам несколько соседних нейронов развивают потенциал действия практически одновременно . Ответ постсинаптического нейрона имеет тот же знак, что и изменения в пресинаптическом нейроне. Так, деполяризация пресинаптической мембраны всегда вызовет деполяризацию постсинаптической мембраны, то же самое имеет место для гиперполяризации . Как правило, ответ постсинаптического нейрона меньше, чем амплитуда исходного сигнала; это обусловлено сопротивлением пре- и постсинаптической мембран. Электрические синапсы имеют сравнительно низкую утомляемость и очень устойчивы к изменениям внешней и внутренней среды . Обычно сигнал может проходить через электрические синапсы в обоих направлениях, однако из этого правила есть исключения. Иногда в ответ на деполяризацию в мембране аксона открываются потенциалзависимые ионные каналы , которые не дают сигналу распространяться в обоих направлениях . Имеются свидетельства своего рода «пластичности» электрических синапсов, то есть электрическая связь между двумя нейронами может ослабляться или усиливаться в зависимости от активности синапса или при изменении внутриклеточной концентрации магния .
Распространение
Электрические синапсы встречаются по всей центральной нервной системе . Они были детально изучены в неокортексе , гиппокампе , , голубом пятне , , , обонятельных луковицах , сетчатке и спинном мозге позвоночных. Электрические синапсы были обнаружены также в полосатых телах , мозжечке и супрахиазматическом ядре . Как правило, электрические синапсы опосредуют очень быстрые поведенческие ответы, как, например, выпускание чернил морским зайцем Aplysia при опасности. Электрические синапсы были обнаружены и вне нервной системы. Электрические синапсы связывают , кардиомиоциты , гладкомышечные клетки , клетки печени , глиальные и эпителиальные клетки .
Электрические синапсы наиболее характерны для низкоорганизованных животных. В ходе эволюции доля электрических синапсов уменьшалась, и в ЦНС млекопитающих (в том числе человека) на долю электрических синапсов приходится около 1 % связей между нейронами .
Клиническое значение
Генетические дефекты коннексинов нередко являются причиной пороков сердца , так как электрические синапсы играют важнейшую роль в синхронизации электрической и сократительной активности сердца . Нарушения в работе коннексинов в шванновских клетках приводят к функциональной патологии аксонов, что лежит в основе болезни Шарко — Мари — Тута . При этом заболевании наблюдается прогрессивная двигательная и сенсорная , кроме того, скорость проведения потенциала действия по аксонам снижена. Образование щелевых контактов между гладкомышечными клетками стенки матки находится под влиянием эстрогенов , которые стимулируют их образование в период беременности . Дефекты щелевых контактов в матке и снижение их числа нередко приводят к преждевременным родам .
История изучения
Впервые гипотеза о существовании электрических синапсов была выдвинута в начале XX века Камилло Гольджи и Йозефом Герлахом . Однако после открытия химических синапсов существование электрических синапсов считалось недостоверным, и до середины XX века главенствовало мнение, что передача потенциала действия между нейронами осуществляется исключительно посредством химических синапсов. Тем не менее, в 1959 году Дэвид Поттер и Эдвин Фершпан убедительно доказали существование электрических синапсов на примере гигантского аксона и аксоном моторного нейрона в брюшной нервной цепочке рака .
Примечания
- ↑ , с. 140.
- ↑ Hormuzdi S. G. , Filippov M. A. , Mitropoulou G. , Monyer H. , Bruzzone R. (англ.) // Biochimica Et Biophysica Acta. — 2004. — 23 March (vol. 1662 , no. 1-2). — P. 113—137 . — doi : . — . [ ]
- Purves, Dale; George J. Augustine; David Fitzpatrick; William C. Hall; Anthony-Samuel LaMantia; James O. McNamara; Leonard E. White. (неопр.) . — 4th. — (англ.) (, 2008. — С. —88. — ISBN 978-0-87893-697-7 .
- ↑ Gibson J. R. , Beierlein M. , Connors B. W. (англ.) // Journal Of Neurophysiology. — 2005. — January (vol. 93 , no. 1). — P. 467—480 . — doi : . — . [ ]
- ↑ Bennett M. V. , Zukin R. S. // Neuron. — 2004. — Vol. 41, № 4. — P. 495—511. — doi : . — . [ ]
- , p. 178–180.
- , p. 178.
- Seung H. Sebastian , Lee Daniel D. , Reis Ben Y. , Tank David W. (англ.) // Journal of Computational Neuroscience. — 2000. — Vol. 9 , no. 2 . — P. 171—185 . — ISSN . — doi : . [ ]
- ↑ , p. 180.
- , с. 146.
- Haas J. S. , Zavala B. , Landisman C. E. (англ.) // Science (New York, N.Y.). — 2011. — 21 October (vol. 334 , no. 6054). — P. 389—393 . — doi : . — . [ ]
- Palacios-Prado N. , Hoge G. , Marandykina A. , Rimkute L. , Chapuis S. , Paulauskas N. , Skeberdis V. A. , O'Brien J. , Pereda A. E. , Bennett M. V. L. , Bukauskas F. F. (англ.) // Journal of Neuroscience. — 2013. — 13 March (vol. 33 , no. 11). — P. 4741—4753 . — ISSN . — doi : . [ ]
- Eugenin E. A. , Basilio D. , Sáez J. C. , Orellana J. A. , Raine C. S. , Bukauskas F. , Bennett M. V. , Berman J. W. (англ.) // Journal Of Neuroimmune Pharmacology : The Official Journal Of The Society On NeuroImmune Pharmacology. — 2012. — September (vol. 7 , no. 3). — P. 499—518 . — doi : . — . [ ]
- Pereda A. E. , Curti S. , Hoge G. , Cachope R. , Flores C. E. , Rash J. E. (англ.) // Biochimica Et Biophysica Acta. — 2013. — January (vol. 1828 , no. 1). — P. 134—146 . — doi : . — . [ ]
- , с. 147—148.
Литература
- Балезина О. П., Гайдуков А. Е., Сергеев И. Ю. Физиология: биопотенциалы и электрическая активность клеток. — М. : Юрайт, 2017. — 165 с. — ISBN 978-5-534-04264-1 .
- Kandel, ER; Schwartz, JH; Jessell, T. M. Principles of Neural Science (неопр.) . — 4th. — New York: McGraw-Hill Education , 2000. — ISBN 0-8385-7701-6 .
- Andrew L. Harris and Darren Locke. (неопр.) . — New York: Springer, 2009. — С. 574. — ISBN 978-1-934115-46-6 .
- Hestrin S. (англ.) // Science (New York, N.Y.). — 2011. — 21 October (vol. 334 , no. 6054). — P. 315—316 . — doi : . — . [ ]
- 2021-03-15
- 2