Слизевики
, или
миксомицеты
(
лат.
Myxomycetes), — класс
настоящих слизевиков
(
Mycetozoa
), включающий более 800 видов организмов, крупнейшая группа среди
амёбозоев
. Встречаются повсеместно
от тропиков до полярных регионов
.
История изучения
Первые зафиксированные наблюдения миксомицетов относятся к XVII веку. Томас Панковиус упоминает
ликогалу древесинную
в своём труде «Herbarium Portatile, oder behendes Kräuter- und Gewächsbuch» 1654 года как «быстрорастущий гриб» (
лат.
Fungus cito crescentes)
. Из-за сходства с
гастеромицетами
миксомицеты в
Systema mycologicum
были отнесены
Элиасом Магнусом Фрисом
к подклассу Myxogastres класса Gasteromycetes. В 1833 году
Генрих Фридрих Линк
выделил миксомицетов в отдельный класс грибов, дав ему название Myxomycetes
.
В 1859 году в своей работе «Die Mycetozoen»
Генрих Антон де Бари
показал отличия миксомицетов от грибов и ввёл термин Mycetozoa («грибо-животные»). В настоящее время установлено, что миксомицеты относятся к группе Amoebozoa, в равной степени близкой грибам и животным. Тем не менее, их изучением по традиции занимаются преимущественно микологи
.
Жизненный цикл и строение
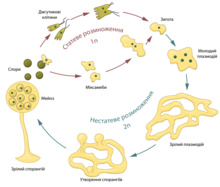
Жизненный цикл миксомицетов
Вышедшие из спор миксамёбы
Плазмодий, формирующий плодовое тело
Жизненный цикл миксомицетов начинается с
гаплоидных
спор
. Из них, в зависимости от влажности окружающей среды, формируются или жгутиковые
зооспоры
, или амёбоидные . Промежуточную стадию между ними принято называть мастигамёбами. При неблагоприятных условиях зооспоры и миксамёбы способны переходить в состояние покоя, образуя микроцисту.
В результате слияния двух гаплоидных клеток образуются диплоидные клетки. Из них путём множественных синхронизированных
митозов
развивается плазмодий — многоядерная, сложно дифференцированная клетка, покрытая только
плазматической мембраной
. Плазмодий обладает отрицательным
фототаксисом
и положительными
гидро-
,
рео-
и , то есть стремится к затенённым влажным местам с большим количеством питательных веществ. При неблагоприятных условиях плазмодии могут формировать покоящиеся стадии — и/или
склероции
.
При определённых условиях плазмодии переходят к формированию спороношений. Все таксисы меняются на обратные. В результате получаются
спорофоры
, которые содержат гаплоидные споры, образовавшиеся посредством
мейоза
. На этом жизненный цикл замыкается
.
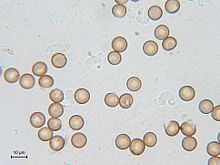
Споры
После разрушения спорофора споры претерпевают период покоя, который иногда может длиться несколько десятков лет
. Споры у большинства миксомицетов сферические, реже эллипсоидальные и яйцевидные. Цвет споровой массы и отдельных спор в проходящем свете — важные диагностические признаки. Оболочка спор может быть снабжена такими структурами как бородавочки (с закруглённой вершиной) и шипы (с заострённой вершиной), но бывают и гладкие споры
.
Миксамёбы и зооспоры
Миксамёбы лишены жёсткой клеточной стенки и способны перемещаться по субстрату с помощью
псевдоподий
. При повышенной влажности они способны за 1,5—2 часа переходить в стадию зооспоры и обратно. В условиях уменьшения влажности и недостатка питания миксамёбы и зооспоры могут образовывать микроцисты с плотными полисахаридными оболочками. При наступлении благоприятных условий микроцисты могут прорасти даже после длительного замораживания и высушивания. При этом оболочка растрескивается и из неё выходят новые миксамёбы или зооспоры
.
Способность к образованию плазмодиев является уникальной чертой миксомицетов. Они представляют собой многоядерную трофическую стадию, способную к амёбоидному движению. Скорость плазмодия может достигать 0,1—0,4 мм в минуту. В плазмодии наблюдаются ритмичные токи цитоплазмы
.
Различают три основных морфологических типа плазмодиев
:
Протоплазмодий
— имеет микроскопические размеры. В нём отсутствуют ритмические токи цитоплазмы. Встречается у представителе порядков Echinosteliales и Cribrariales.
Афаноплазмодий
— характеризуется отсутствием развитого слизевого чехла и наличием особой стадии «коралла» перед началом формирования спорофоров. Характерен для представителей порядка Stemonitidales. Наиболее устойчив к обезвоживанию
.
Фанероплазмодий
— обладает слизистым чехлом и ритмичными токами цитоплазмы. Часто бывает ярко окрашен. Наиболее характерен у представителей порядка Physarales, но имеется и у некоторых представителей Cribrariales.
Иногда выделяют ещё один тип плазмодия, занимающий по морфологии промежуточное положение между афаноплазмодием и фанероплазмодием. Он характеризуется наличием токов цитоплазмы, но имеет меньшие размеры и более слабую пигментацию. Такой тип плазмодия характерен для представителей порядка Trichiales
.
В неблагоприятных условиях плазмодии могут образовывать склероции или фрагментироваться на отдельные сгустки цитоплазмы, покрытые плотной оболочкой — сферулы
.
Спороношения
Выделяют четыре основных типа спороношений миксомицетов, между которыми бывают промежуточные варианты
:
Плазмодиокарпы
— наиболее простой тип спорофоров. Образуется при созревании плазмодия, покрытого общей оболочкой, почти без изменения его конфигурации. По форме представляется собой короткие или длинные шнуры, вытянутые ветвящиеся или сетчатые образования.
Спорангии
— шаровидные, овальные, колончатые или другие спороносные структуры, формирующиеся в результате дробления плазмодия. Могут быть сидячими или на ножках.
Псевдоэталии
образуются при срастании спорангиев боковыми стенками и потере ими индивидуальности, с сохранением возможности подсчитать количество спорангиев.
Эталии
— тип спорофоров, образующийся при срастании спорангиев с полной потерей их индивидуальности. Представляют собой подушковидные образования, покрытые общей оболочкой (кортексом).
Трофические стадии миксомицетов, такие как миксамёбы, представляющие собой доминирующую группу почвенных
простейших
, питаются бактериями
. Бо́льшую часть информации об экологии этих организмов получают при изучении их спорофоров, из-за чего исследователи в основном имеют дело с данными об их расселительных стадиях
.
На основании изучения особенностей распространения спорофоров выделяют следующие эколого-трофические группы
:
Лигнофильные
— встречаются на гниющей древесине. Самая большая группа, включающая до 70 % известных миксомицетов. Большинство из них обладают макроскопическими размерами. Пики спороношений наблюдаются с середины лета до поздней осени.
Кортикальные
— обитают на коре живых деревьев и кустарников. Обычно небольшие (от 100 мкм до 1—2 мм). Доминируют в пустынях и местах с высокой антропогенной нагрузкой.
Подстилочные и почвенные
— питаются в верхнем почвенном слое. Спороношения образуют на растительных остатках. Видовой состав в разных слоях листового опада неоднороден
.
Копрофильные
— обитают на выветренном
помёте
растительноядных
млекопитающих
и
птиц
. Наибольшего разнообразия достигают в зонах
аридного климата
. За всё время исследования было обнаружено 114 видов копрофильных миксомицетов, 16 из которых были специфичны для данного типа субстрата
. Представителей этой группы редко находят в полевых условиях. Основным методом их изучения является метод «влажной камеры». В отличие от грибов, большинство копрофильных миксомицетов не проходят через
желудочно-кишечный тракт
животных, а заселяют субстрат спустя продолжительное время. В то же время, споры некоторых видов обладают толстыми стенками, что может быть адаптацией к прохождению через пищеварительную систему
.
Бриофильные
— обитают на
мхах
. Часто ассоциированы с микроскопическими
водорослями
. Являются самой малочисленной группой, включающей менее 5 % известных видов. Большинство представителей имеют микроскопические размеры. Появляются поздней осенью.
Нивальные
— развиваются весной на растительных остатках возле тающего снега. Наибольшего разнообразия достигают в альпийских и субальпийских районах северного полушария
, но могут встречаться и в горах южного полушария
и на равнинах
. Для развития спороношений нивальных миксомицетов необходимо наличие снежного покрова в осенние месяце для предотвращения промерзания почвы
.
Один и тот же вид миксомицетов может обитать на разных типах субстрата и потому может быть отнесён к разным эколого-трофическим группам.
Таксономия
Классическая система
Класс
Myxomycetes
включает пять порядков.
Echinosteliales
Порядок
включает 2 семейства —
и
. Эхиностелиевые рассматриваются как наиболее близкая к группа миксомицетов. Это заключение носит предварительный характер, так как ультраструктура жгутиков зооспор некоторых протостелиевых и видов рода
отличается. В то же время наблюдается значительное сходство в строении одноядерных миксамёб протостелиевых и протоплазмодия эхиностелиевых, а также сходство морфологии плодовых тел у представителей обеих групп. Очевидно, для выяснения положения рода
требуются дополнительные молекулярно-биологические исследования.
Порядок
включает 3 семейства:
,
,
, для представителей которых характерно отсутствие
капиллиция
. Однако этот признак встречается также у рода
из порядка
. Некоторые виды рода
обладают сходными признаками с
(
Trichiales
) и
(
Liceales
), что ставит под сомнение гомогенность рода в современном понимании его объёма.
Trichiales
Порядок
включает 2 семейства:
и
Trichiaceae
. Границы между родами не всегда чётко определены, так же как и их положение в системе. У видов первого нити капиллиция сплошные, у второго — полые. В пределах этого семейства усложнение капиллиция происходило как за счёт усложнения системы капиллиция в целом, так и её отдельных элементов. Наиболее просто устроен капиллиций у представителей рода
Perichaena
, состоящий из редко ветвящихся трубочек. Очевидно, такой тип можно рассматривать как плезиоморфный признак. Отсутствие чётко выраженного протока в нитях капиллиция у видов рода
, возможно, потребует пересмотра положения этого рода в семействе
Trichiaceae
. Следующим этапом усложнения капиллиция считается появление почти неорнаментированных и не связанных с
перидием
элатер, как у представителей рода
. У видов рода
заметно усложнение орнаментации нитей капиллиция за счёт образования на них шипиков и спиральных утолщений. Связь капиллиция с перидием и образование сети нитей, а также их способность вытягиваться при созревании спорангия и изменении влажности, что приводит к растрескиванию перидия, является следующим этапом дифференциации этой структуры. Подобный тип характерен для родов
и
. Основное отличие между ними заключается в различной орнаментации отдельных трубочек капиллиция.
Порядок
включает два семейства
и
. На основании наблюдений за развитием и в результате электронно-микроскопических исследований роды
,
,
были перемещены из
Stemonitales
в
Physarales
. Что касается родов
Diachea
и
Leptoderma
, то развитие спорофора по субгипоталлическому типу и наличие у видов этих родов указывают на их принадлежность к порядку
Physarales
.
Подкласс
Lucisporomycetidae
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin — включает представителей со светлыми спорами без
меланина
.
Надпорядок
Cribrariidia
Порядок
Cribrariales
T. Macbr.
Семейство
Cribrariaceae
Corda
Надпорядок
Trichiidia
Порядок
Liceales
E. Jahn
Семейство
Liceaceae
Chevall.
Порядок
Reticulariales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
Семейство
Reticulariaceae
Chevall.
Порядок
Trichiales
Chevall.
Семейство
Trichiaceae
Chevall.
Семейство
Dianemataceae
T. Macbr.
Lucisporomycetidae
incertae sedis (светлоспоровые роды с неясным систематическим положением)
Hochg. & Gottsb.
Morgan
G. Lister
Novozh., Hoof & Jagers
Подкласс
Columellomycetidae
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin — включает преимущественно представителей с тёмной споровой массой, окраска которой определяется содержанием меланина.
Надпорядок
Echinosteliidia
Порядок
Echinosteliales
G. W. Martin
Семейство
Echinosteliacea
Rostaf. ex Cooke
Надпорядок
Stemonitidia
Порядок
Clastodermales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
Семейство
Clastodermataceae
Alexop. & T. E. Brooks
Порядок
Meridermatales
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
Семейство
Leontyev, Schnittler, S.L. Stephenson, Novozhilov & Shchepin
Порядок
Physarales
T. Macbr.
Семейство
Didymiaceae
Rostaf. ex Cooke
Семейство
T. Macbr.
Семейство
Physaraceae
Chevall.
Порядок
Stemonitidales
T. Macbr.
Семейство
Rostaf. ex Cooke
Семейство
Stemonitidaceae
Fr.
Columellomycetidae
incertae sedis (темноспоровые роды с неясным систематическим положением)
Fr.
Reinhardt & L. S. Olive
G. Lister
Hertel
M. Blackw. & Alexop.
H. W. Keller
Kuntze
Примечания
(англ.)
на сайте
MycoBank
.
(Дата обращения: 18 февраля 2020)
.
↑ , с. 16—20.
Steven L. Stephenson, Yuri K. Novozhilov, Martin Schnittler.
(англ.)
// Journal of Biogeography. — 2000-05. — Vol. 27 , iss. 3 . — P. 741–754 . —
ISSN
. —
doi
: .
Steven L. Stephenson, Rodney D. Seppelt, Gary A. Laursen.
(англ.)
// Antarctic Science. — 1992-12. — Vol. 4 , iss. 4 . — P. 431–432 . —
ISSN
. —
doi
: .
↑
Dmitry V. Leontyev, Martin Schnittler, Steven L. Stephenson, Yuri K. Novozhilov, Oleg N. Shchepin.
// Phytotaxa. — 2019-03-27. — Т. 399 , вып. 3 . — С. 209 . —
ISSN
. —
doi
: .
13 января 2022 года.
, с. 11.
, с. 5.
↑ , с. 7—15.
Ernest C. Smith.
// Mycologia. — 1929-11. — Т. 21 , вып. 6 . — С. 321 . —
doi
: .
1 января 2022 года.
Eugene W. Elliott.
(англ.)
// Mycologia. — 1949-03. — Vol. 41 , iss. 2 . — P. 141–170 . —
ISSN
. —
doi
: .
, с. 23—25.
Collins O. R.
(англ.)
// Botanical Review. — 1979. — Vol. 45 , no. 2 . — P. 145-201 .
1 января 2022 года.
Steven L. Stephenson, Anna Maria Fiore-Donno, Martin Schnittler.
(англ.)
// Soil Biology and Biochemistry. — 2011-11. — Vol. 43 , iss. 11 . — P. 2237–2242 . —
doi
: .
30 июля 2020 года.
M.F. Madelin.
(англ.)
// Transactions of the British Mycological Society. — 1984-08. — Vol. 83 , iss. 1 . — P. IN1–19 . —
doi
: .
25 июня 2018 года.
Aw Rollins.
// Mycosphere. — 2012-09-08. — Т. 3 , вып. 5 . — С. 543–549 . —
doi
: .
1 января 2022 года.
↑
Uno Eliasson.
(англ.)
// Fungal Diversity. — 2013-03. — Vol. 59 , iss. 1 . — P. 85–90 . —
ISSN
. —
doi
: .
Y. K. Novozhilov, M. Schnittler, D. A. Erastova, M. V. Okun, O. N. Schepin.
(англ.)
// Fungal Diversity. — 2013-03. — Vol. 59 , iss. 1 . — P. 109–130 . —
ISSN
. —
doi
: .
Steven L. Stephenson, John D. L. Shadwick.
(англ.)
// Australian Journal of Botany. — 2009. — Vol. 57 , iss. 2 . — P. 116 . —
ISSN
. —
doi
: .
Ерастова Д. А. , Новожилов Ю. К.
// Микология и фитопатология. — 15. — Т. 49 , № 1 . — С. 9—18 . —
ISSN
.
, с. 347—353.
Литература
Гмошинский В. И. , Дунаев Е. А. , Киреева Н. И.
Определитель миксомицетов Московского региона. —
М.
: АРХЭ, 2021. — 388 с. —
ISBN 978-5-94193-089-0
.
Новожилов Ю. К.
Определитель грибов России: Отдел Слизевики. —
СПб.
: Наука, 1993. — 288 с. —
ISBN 5-02-026625-0
.
Fiore-Donno A.M., Berney C., Pawlowski J., Baldauf S.L.
higher-order phylogeny of plasmodial slime molds (Myxogastria) based on elongation factor 1-A and small subunit rRNA gene sequences
(англ.)
// J. Eukaryot. Microbiol. : journal. — 2005. — Vol. 52 , no. 3 . — P. 201—210 . —
doi
: . — .
Kirk P. M., Cannon P. F., Minter D. W., Stalpers JA.
(неопр.)
. — Wallingford: CABI, 2008. — С. . —
ISBN 0-85199-826-7
.
Baldauf S.L., Doolittle W.F.
(англ.)
// Proc. Natl. Acad. Sci. U.S.A. : journal. — 1997. — October (vol. 94 , no. 22). — P. 12007—12012 . —
doi
: . — . —
PMC
.
Stephenson S. L. , Rojas C.
Myxomycetes: biology, systematics, biogeography and ecology
(англ.)
. — 2nd ed. — Elsevier: Academic Press, 2021. — 600 p. —
ISBN 9780128242827
.