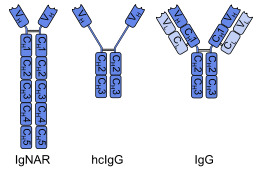
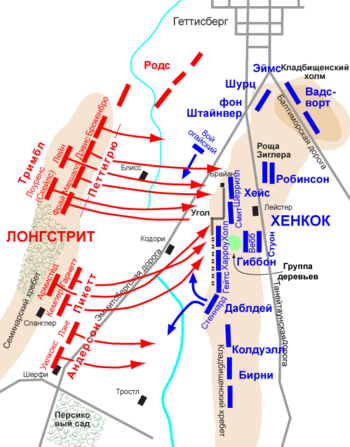
Молекулы
антител
состоят из двух тяжёлых и двух
лёгких
цепей. Различные варианты тяжёлых цепей соответствуют
изотипам иммуноглобулинов
. Типы тяжёлых цепей отличаются у разных видов животных. Все тяжёлые цепи содержат иммуноглобулиновые
домены
— обычно один вариабельный домен, который обуславливает связывание антигена и несколько константных доменов.
У млекопитающих
Классы
У
млекопитающих
классифицируют пять типов тяжёлых цепей γ, δ, α, μ и ε.
Типы тяжёлых цепей соответствуют классам иммуноглобулинов.
Тяжёлые цепи α и γ состоят примерно из 450 остатков аминокислот.
Тяжёлые цепи μ и ε состоят примерно из 550 остатков аминокислот
.
Участки
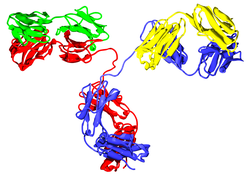
Две тяжёлых цепи (показаны синим) соединены дисульфидными связями с двумя лёгкими цепями (показаны зелёным). Показаны константные и вариабельные домены.
Каждая тяжелая цепь имеет два участка:
константный участок, одинаковый у всех иммуноглобулинов одного класса, но отличающийся между классами
тяжёлые цепи γ, α и δ имеют константный участок, состоящий из трёх тандемных иммуноглобулиновых доменов, а также шарнирный участок, обеспечивающий гибкость молекулы.
тяжёлые цепи μ и ε имеют константный участок, состоящий из четырёх иммуноглобулиновых доменов
.
вариабельный участок, который отличается между разными В-лимфоцитами, но одинаковый среди всех иммуноглобулинов, синтезированных одним В-лимфоцитом или одним клоном. Вариабельные домены всех тяжёлых цепей состоят из одного иммуноглобулинового домена. Такие домены имеют длину около 110 остатков аминокислот.
У рыб
являются наиболее примитивными животными, способными образовывать антитела, подобные тем, что образуют млекопитающие.
Однако, рыбы не имеют такого репертуара антител, какой характерен для млекопитающих.
Три различных вида тяжёлых цепей обнаружены и у костных рыб.
Первыми описанными тяжёлыми цепями были μ (или
мю
). Тяжёлые цепи такого типа представлены у всех челюстных рыб и считаются базовыми для всех иммуноглобулинов. Образующиеся антитела IgM, секретируются у челюстных рыб в виде
тетрамера
(в отличие от акул и млекопитающих, у которых IgM представляют собой).
Тяжёлые цепи δ, представленные в IgD были впервые обнаружены у сомов и
атлантического лосося
, и в настоящее время описаны и для других костистых рыб.
Третий тип тяжёлых цепей костистых рыб описан относительно недавно, и не соответствует какому-либо типу тяжёлых цепей млекопитающих. Такие тяжёлые цепи описаны для
радужной форели
(τ)
и
Данио рерио
(ζ).
Три различных типа тяжёлых цепей иммуноглобулинов описаны и у
хрящевых рыб
.
Примечания
↑
Janeway C. A., Jr.
et al.
Immunobiology
(неопр.)
. — 5th ed.. —
(англ.)
(, 2001.
Woof J., Burton D.
Human antibody-Fc receptor interactions illuminated by crystal structures
(англ.)
//
Nat Rev Immunol
: journal. —
Nature Publishing Group
, 2004. — Vol. 4 , no. 2 . — P. 89—99 . —
doi
: . — .
23 марта 2007 года.
Eva Bengtén, L. William Clem, Norman W. Miller, Gregory W. Warr and Melanie Wilson.
Channel catfish immunoglobulins: Repertoire and expression
. Developmental & Comparative Immunology, Volume 30, Issues 1-2, Antibody repertoire development, 2006, Pages 77-92.
Stein Tore Solem and Jørgen Stenvik.
Antibody repertoire development in teleosts--a review with emphasis on salmonids and Gadus morhua L
. Developmental & Comparative Immunology, Volume 30, Issues 1-2, Antibody repertoire development, 2006, Pages 57-76.
J.D. Hansen, E.D. Landis and R.B. Phillips.
Discovery of a unique Ig heavy-chain isotype (IgT) in rainbow trout: Implications for a distinctive B cell developmental pathway in teleost fish.
Proceedings of the National Academy of Sciences U S A. Volume 102, Issue 19, 2005, pages 6919-24.
N. Danilova, J. Bussmann, K. Jekosch, L.A Steiner.
The immunoglobulin heavy-chain locus in zebrafish: identification and expression of a previously unknown isotype, immunoglobulin Z.
Nature Immunology, Volume 6, Issue 3, 2005, pages 295—302.
H. Dooley and M.F. Flajnik.
Antibody repertoire development in cartilaginous fish
. Developmental & Comparative Immunology, Volume 30, Issues 1-2, Antibody repertoire development, 2006, Pages 43-56.