Кладистика
- 1 year ago
- 0
- 0


Клади́стика (от др.-греч. κλάδος [kládos] «ветвь») — направление филогенетической систематики . Характерные особенности кладистической практики состоят в использовании так называемого кладистического анализа (строгой схемы аргументации при реконструкции родственных отношений между таксонами ), строгом понимании монофилии и требовании взаимно-однозначного соответствия между реконструированной филогенией и иерархической классификацией [ источник не указан 2176 дней ] . Кладистический анализ — основа большинства принятых в настоящее время биологических классификаций, построенных с учётом родственных отношений между живыми организмами [ источник не указан 2176 дней ] .
Кладистика относится к числу трёх ведущих таксономических школ , доминирующих в современной биологической систематике ; ей противостоят фенетика , основанная на количественной оценке так называемого общего сходства ( англ. overall similarity ), и эволюционная таксономия , которая, подобно кладистике, при построении системы опирается на эволюционную близость (то есть общность происхождения), однако не требует строгого соответствия системы и филогении (в частности, это выражается в признании права на существование в системе парафилетических групп) .
Кладистика основана на идеях немецкого энтомолога и систематика Вилли Хеннига (1913—1976), изложенных в его работах 1950—1960-х гг., однако название кладистика было впервые использовано только в середине 1960-х годов критиками этого направления филогенетической систематики, а современные формализованные процедуры были разработаны только в 1970-е гг. его последователями в США.
Уже к концу XX века кладистика распалась на несколько школ, которых фактически объединяет лишь использование нескольких знаковых символов. К их числу относятся эволюционная кладистика ( англ. evolutionary cladistics ), экономная кладистика ( англ. parsimony cladistics ) и структурная кладистика (или паттерн-кладистика ) .
Классификационный подход, который сейчас известен как кладистика , был заложен работами Вилли Хеннига . Но его главная книга «Grundzüge einer Theorie der Phylogenetischen Systematik» не содержит (даже в редакции 1979 года ) термина «кладистика», и автор книги ссылается на свой собственный подход как на «филогенетическую систематику». в своей обзорной статье утверждает, что термин « клада » был предложен в 1958 году Джулианом Хаксли , «кладистика» — и Дж. Харрисоном в 1960 году , а «кладист» (для обозначения последователя школы Хеннига) — Э. Майром в 1965 году .
Некоторые дебаты, в которые были вовлечены кладисты, велись ещё XIX века, но кладисты присоединились к ним позже и с новыми взглядами, что описано в предисловии к книге Хеннига редакции 1979 года, где Розен, Нельсон и Патерсон, нападая на методологические основы эволюционной таксономии, отмечали следующее:
Отягощённый неясными и скользкими идеями об адаптации, эволюционной пригодности, биологических видах и естественном отборе, неодарвинизм (который определяется как «эволюционный» подход к биологической систематике Майром и Симпсоном) не имеет определенного метода исследования, а также пришёл в зависимость от консенсуса авторитетов по вопросам эволюционной интерпретации и классификации.

Апоморфный признак (или апоморфия ) — это характеристический признак, который отличается от унаследованного членами группы признака их общего предка и ставший уникальным, выделяющий группу видов среди множества других.
Плезиоморфный признак (или плезиоморфия ) — это характеристический признак, унаследованный видами группы видов от их общего предка и характеризующий бо́льшую по объёму группу, содержащую первую в качестве своей подгруппы . Прилагательные «плезиоморфный» и «апоморфный» используются вместо «примитивный» и «продвинутый» для предотвращения оценочного суждения об эволюционных формах, поскольку оба они могут обозначать структуры, крайне полезные при некоторых обстоятельствах и вредные — при других. Нередко набор плезиоморфных форм неформально называется «базисом» для клады или клад, которым этот набор присущ.
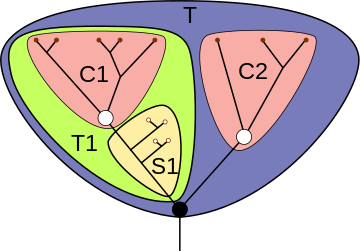
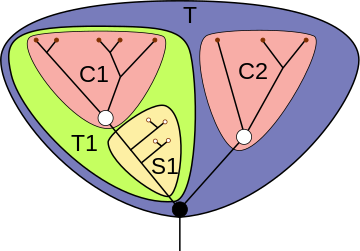
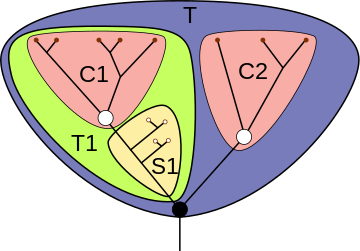
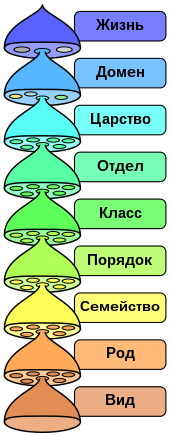
Для наглядного представления филогенетических взаимоотношений между различными таксонами в кладистике принято использовать кладограммы — древовидные схемы предполагаемых генеалогических связей таксонов различного уровня, отображающие иерархию монофилетических групп . При этом терминальным таксонам отвечают терминальные элементы кладограммы, нетерминальным — её рёбра (то есть междоузлия — вообще говоря, не все, если не всякая клада получает в данной системе статус таксона). Узловые точки кладограммы не соотнесены никаким таксонам: они соответствуют анцестральным (предковым) видам отдельных нетерминальных таксонов, причём такие виды всегда трактуются как гипотетические и не специализированы .
Ряд распространённых в кладистике терминов введён для описания позиций таксонов внутри кладограммы. Вид или клада является базальной группой относительно другой клады, если первая имеет больше плезиоморфных черт, чем вторая. Базальная группа, как правило, насчитывает меньшее количество видов по сравнению с развитыми группами; наличие в кладограмме базальной группы не является обязательным.
Клада или вид, находящийся в кладограмме внутри другой клады, может называться «вложенной» или «вложенным» в эту кладу.

Массив информации, который может быть подвергнут кладистическому анализу, должен быть организован специальным образом. Для этого, прежде всего, необходимо провести разграничение между признаками и их состояниями . Например, цвет перьев может быть голубым у одного вида птиц и красным у другого. Тогда «голубые перья» и «красные перья» будут двумя состояниями одного признака — «цвета перьев».
После этого устанавливают полярность признаков: у каждого из них вычленяются два состояния — примитивное и продвинутое. Это делают на основании сравнения исследуемых групп с внешней группой ( англ. outgroup ) — таксоном, не входящим в исследуемую совокупность, но филогенетически близким им. Примитивным состоянием при этом признаётся такое состояние признака, которое наблюдается и у внешней группы .
В дальнейшем примитивное состояние признака может быть интерпретировано как плезиоморфия, а продвинутое — как апоморфия. Апоморфия, присущая данным двум группам (или большему их числу) называется синапоморфией ; предполагается, что она имелась и у последнего общего предшественника данных групп, но до его появления отсутствовала. Надо заметить, что при выделении клад могут использоваться только синапоморфии .
Следующей стадией является составление различных возможных кладограмм и их проверка. В идеале клады имеют много «согласованных» синапоморфий; в таком идеальном случае ожидают наличие достаточно большого количества настоящих синапоморфий, которые не могут быть скрытыми гомоплазиями , появление которых вызывает конвергентная эволюция (то есть качествами, которые воспроизводят друг друга благодаря влиянию окружающей среды или общего функционального использования, а не всеобщему происхождению). Известным примером гомоплазии, возникшей благодаря конвергентной эволюции, служит признак «наличие крыльев». Хотя крылья птиц , летучих мышей и насекомых выполняют одинаковые функции, каждое из этих крыльев эволюционировало независимо, что может быть прослежено благодаря их анатомии. Если же птица, мышь и крылатое насекомое будут объединены признаком «наличие крыльев», в массив данных будет искусственно внесена гомоплазия, что разрушит анализ и скорее всего приведёт в результате к неверному построению эволюционной картины.
Применение гомоплазии в морфологических наборах данных часто можно избежать путём точнейшего определения характерных признаков и увеличения их количества: в предыдущем примере, используя в качестве признаков «крылья с перьями», «крылья с хитиновым экзоскелетом» и «кожистые крылья» как характерные признаки, можно избежать эволюционно ложного объединения трех перечисленных групп животных на основе гомоплазии. При анализе «супердеревьев» (баз данных, включающих большее число таксонов исследуемой клады), применение неточных признаков может стать неизбежным, поскольку в обратном случае признаки могут стать неприменимыми для всех многочисленных таксонов. Скажем, такой признак, как наличие крыльев, не может быть применен для анализа филогении настоящих многоклеточных животных ( Eumetazoa ), так как большинству видов этого таксона данный признак не присущ. Таким образом, осторожный выбор и определение характерных признаков является другим важным элементом кладистического анализа. При ошибочном определении корневого вида и набора признаков, никакие методы построения кладограмм не смогут дать в результате филогенетической системы, которая соответствует эволюционной реальности.
Для практически любого таксона можно построить не одну, а множество различных кладограмм, основываясь на различных корневых видах и наборах характерных признаков; но из них выбирают одну-единственную, руководствуясь принципом парсимонии : компактную систему, которая, в сочетании с наименьшими возможными изменениями характерных признаков (синаптоморф), даёт непротиворечивую картину происхождения клады (в общем, это вариант соображения по бритве Оккама ). Хотя в начале такой анализ проводился «вручную», впоследствии для него стали применяться компьютеры со специальным программным обеспечением, которое позволяет оперировать на порядки большими наборами данных и количеством признаков. Такие программы (вроде и других подобных) позволяют делать статистическую оценку вероятности нод (узлов и разветвлений) построенной кладограммы.
Важно также заметить, что ноды кладограммы не обязательно отражают различия эволюционных ветвей, а лишь различия постоянных признаков, которые наблюдаются между этими ветвями. Признаки, которые заключаются в разнице последовательностей ДНК , способны расходиться после того, как генный дрейф между популяциями редуцируется к некоторой пороговой величине, в то время как заметные морфологические изменения, обычно будучи эпистатическими (то есть результатом взаимодействия нескольких генов) выявляются только после того, как разошедшиеся таксоны отдельно эволюционно развивались в течение некоторого (обычно довольно значительного) времени; так, биологические подвиды зачастую могут быть разрознены генетически, но не морфологически (по строению тела или внутренней анатомии).
Учитывая то, что секвенирование ДНК становится всё дешевле и проще во внедрении, молекулярная систематика приобретает все большую популярность как способ реконструкции филогенеза . Использование критерия парсимонии является лишь одним из методов нахождения филогении таксона на базе молекулярно-биологических данных; методы максимального правдоподобия и Байесовского вывода , которые включают в себя детально разработанные модели эволюции последовательностей, являются известными не- хеннигскими методами оценки данных секвенирования. Другим мощным методом реконструкции филогении является использование ретротранспозонных маркеров генома, которые считаются таковыми, потому что менее подвержены воздействию реверсии секвенированных данных, которая (реверсия) может сделать данные непригодными для построения филогенетической последовательности. Также преимуществом ретротранспозонных маркеров является то, что они мало подвержены гомоплазии из-за интегрирования в геном в полностью случайных местах (хотя такая статистическая случайность является сомнительной).
В идеале морфологические, молекулярно-биологические и другие (поведенческие, экологические, палеонтологические, и т. д.) филогенетические данные должны обобщаться при разработке итогового заключения, при этом никакой из методов не является более доказательным, чем другие, но все они имеют разные внутренние источники ошибок. Например, конвергенция развития признаков (гомоплазия) гораздо чаще появляется при анализе морфологических данных, чем в данных, полученных при молекулярном секвенировании, но реверсии признаков у них обоих встречаются с примерно одинаковой частотой; обычно, морфологические гомоплазии могут быть вскрыты при достаточно внимательном и детальном анализе характерных признаков.
Кладистика не отдаёт предпочтения ни одной из имеющихся теорий эволюции, принимая во внимание только общую информацию о происхождении с существенными её модификациями. Благодаря этому кладистические методы могут быть (и бывают) использованными в небиологических дисциплинах, включая историческую лингвистику и задачи по определению авторства текстов.

С 1960-х в биологии возникла тенденция под названием «кладизм» или «кладистическая таксономия» считать таксоны кладами. Иными словами, кладисты считают, что биологическая классификация должна быть реформирована таким образом, чтобы устранить все не-клады. В отличие от кладистов, все остальные таксономисты настаивают на том, что каждая из таксономических групп живых существ отражает филогенетическую историю. Последние часто используют кладистический подход, но также допускают в формировании классификационных деревьев использование как монофилетических (что совпадает с кладистичним подходом), так и парафилетических таксонов. Как результат, уже как минимум с начала XX века роды и таксоны низшего уровня формировали основываясь на монофилетическом подходе, в то время как таксоны высокого ранга могут быть (а такие как класс и выше — обычно и являются) парафилетическими.
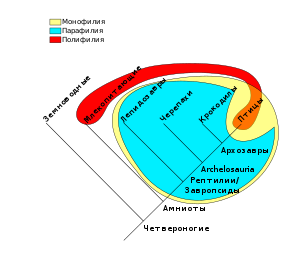
В кладистике монофилетической группой считается клада , состоящая из предшественника и всех его потомков, которые формируют одну и только одну эволюционную группу. Парафилетической является такая же, но без некоторых потомков, которые претерпели существенные изменения. Например, традиционный класс Пресмыкающиеся не включает птиц, хотя птицы произошли от пресмыкающихся; аналогично, традиционная группа Беспозвоночные является парафилетической, потому что не включает позвоночных, которые происходят от неё.
Группа, состоящая из членов, которые происходят из разных эволюционных линий, называется полифилетической . Например, сформированная ранее таксономическая группа «толстокожие» ( ) была затем признана полифилетической, поскольку включенные в неё слоны, носороги и бегемоты произошли от разных, не родственных между собой предшественников. Аналогично, сформированный прежде тип Кишечнополостные был позднее разделён на книдарий и гребневиков на основании их происхождения от различных предшественников и возникших вследствие этого существенных морфологических различий. Эволюционные таксономисты считают полифилетические группы ошибками классификации, вызванными конвергенцией и другими видами гомоплазии, которые ошибочно интерпретируются как гомологии.
Следуя Хеннигу, кладисты считают, что парафилия является не менее неприемлемой для классификации, чем полифилия . В рамках кладистики считается, что монофилетические группы могут быть объективно и строго определёнными, основываясь на общих предшественниках или синаптоморфах. В отличие от них, парафилетические и полифилетические группы выделяются на основе анализа ключевых характеристик, в ходе которого степень важности этих характеристик является весьма субъективной. Кладисты отмечают, что, таким образом, это приводит к «градистическому» мышлению, которое в свою очередь тяготеет к телеологии , которую обычно считают неприемлемой в эволюционных исследованиях, поскольку телеология подразумевает существование общего плана эволюции, который не может быть показан экспериментально.
Продолжая эту линию критики, некоторые кладисты утверждают, что таксоны высшего по виду ранга являются слишком субъективными, и поэтому их выделение не представляет никакой содержательной информации. На основе этого, как утверждают такие критики, эти таксоны не должны использоваться в систематике вообще. Таким образом, биологическая систематика, в их понимании, должна полностью отойти от Линнеевской таксономии и стать простой иерархией клад. Весомость этой системы аргументов критически зависит от того, как часто в ходе эволюции прерывается градуалистическое равновесие экологических условий. Квази-стабильное состояние экосистем приводит к медленному филогенезу, который может быть трудно интерпретирован в рамках линнеевской систематики; в то же время катастрофическое событие выводит таксон из экологического равновесия и обычно приводит к разделению признаков клад. Это разделение происходит в течение эволюционно короткого времени, и в результате дает систему таксонов, которая легко описывается в рамках линнеевской классификации.
Другие эволюционные систематики утверждают, что все таксоны по своей сути субъективны, даже если они отражают эволюционные взаимоотношения, поскольку живые организмы формируют непрерывное эволюционное дерево . В таком случае, любая линия раздела является искусственной и создает монофилетическое выделение над парафилетическим. Парафилетические таксоны являются необходимыми для классификации более ранних секций дерева — например, ранние позвоночные , которые через некоторое время развились в семью Гоминиды , не могут быть помещены более в одну монофилетическую семью. Также в этой системе аргументов приводится то, что парафилетические таксоны предоставляют информацию о существенных изменениях в морфологии, экологии и эволюционной истории организмов — короче говоря, что и таксоны, и клады являются ценными для построения истинной картины систематики живой природы, но и те, и те — с некоторыми ограничениями.
В полемике вокруг годности кладистических методов при построении биологической классификации термин « монофилия » часто используется в своем старом смысле, где он включает в себя парафилию, наряду с чем используется термин «голофилия» для описания «чистых» клад («монофилии» в Хеннигском смысле). Как пример ненаучности эмпирического метода используется следующий: если какая-то парафилетическая эволюционная линия приводит к возникновению клады из-за значительной адаптивной радиации и многочисленных, хорошо заметных и важных синапоморфий, особенно таких, что ранее в этой группе живых существ не встречались, парафилия не признается достаточным аргументом против выделения таксона в линнеевской системе классификации (но это считается достаточным контраргументом в рамках филогенетической номенклатуры). Например, как было указано выше, и завропсиды («рептилии»), и птицы в линнеевской систематике имеют одинаковый ранг классов, хотя последние являются эволюционно крайне специализированной ветвью рептилий.
Формальный сборник филогенетической номенклатуры ( ) сейчас находится в разработке для приспособления его к кладистической таксономии. Его планируют использовать и те, кто пытается полностью избегать линнеевской номенклатуры, и те, кто использует в систематике таксоны вместе с кладами. При некоторых обстоятельствах (например, при классификации гесперорнисов ) методы ФилоКода были привлечены во избежание неопределенностей, которые возникали при применении линнеевской систематики к этому таксону; таким образом, комбинация этих двух систем позволяет сформировать таксономическую картину, которая размещает группу живых существ на эволюционном дереве в полном соответствии с имеющейся научной информацией.