Список губернаторов Небраски
- 1 year ago
- 0
- 0

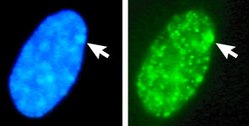
Гетерохромати́н — участки хроматина , находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость .
В 1907 году немецкий цитолог С. Гутхерц ( S. Gutherz ) обнаружил, что некоторые фрагменты хромосом или хромосомы целиком во время клеточного деления интенсивно окрашиваются и выглядят более конденсированными по сравнению со слабоокрашенными участками. Это явление было названо гетеропикнозом, однако термин в дальнейшем не прижился . В ядрах клеток , находящихся в интерфазе , были обнаружены участки, интенсивно окрашиваемые красителями, связывающимися с хроматином, такие участки были названы хромоцентрами. С. Гутхерц показал, что гетеропикнотические сегменты хромосом становятся заметными в начале профазы, то есть в начале конденсации хромосом, отличаясь от «нормальных» участков более интенсивной окраской; различия в интенсивности окраски по мере конденсации снижаются и становятся практически неразличимыми в конце метафазы.
Другой немецкий цитолог , проанализировав соотношение числа хромоцентров и гетеропикнотических участков хромосом, наблюдаемых при митозе в клетках мха, пришёл к выводу, что хромоцентры, обнаруживаемые в интерфазе, ассоциируются с сильно конденсированными и интенсивно окрашиваемыми гетеропикнотическими участками хромосом, наблюдаемыми в течение митотического цикла, то есть хромоцентры и гетеропикнотические участки являются одними и теми же участками хромосом, которые не подвергаются деконденсации в телофазе.
В 1928 году Хайц предложил термины « эухроматин » для участков хромосом, претерпевающих процесс компактизации-декомпактизации в процессе митоза и «гетерохроматин» для участков, постоянно остающихся в конденсированном состоянии. Хайц считал, что гетерохроматиновые участки хромосом являются генетически инертными .
Основное функциональное отличие факультативного гетерохроматина от конститутивного — возможность перехода в эухроматиновое состояние, при котором ДНК становится транскрипционно активной и, соответственно, происходит экспрессия генов , локализованных на данном участке хромосомы.
Факультативный гетерохроматин содержит кодирующую и, в силу этого, относительно консервативную ДНК; ДНК конститутивного гетерохроматина является преимущественно некодирующей и в силу этого высокополиморфна и вариабельна.
На ранних стадиях онтогенеза во многих случаях содержание гетерохроматина в метафазных хромосомах значительно ниже, чем на более поздних стадиях и в клетках взрослого организма — метафазные хромосомы бластомеров многих позвоночных сильно декомпактизированы, в интерфазных ядрах гетерохроматиновые образования не обнаруживаются.
Факультативный и конститутивный гетерохроматины также выявляются по различию в окрашивании: если факультативный гетерохроматин подвергается G-окрашиванию по Романовскому — Гимзе в стандартных условиях, то окраска этим же красителем после -ренатурации ДНК избирательно окрашивает конститутивный гетерохроматин. Такой селективный метод получил название окраски на конститутивный (C) гетерохроматин, или C-окрашивание .
Обычно факультативные гетерохроматиновые участки присутствуют только в одной из гомологичных хромосом. Типичным примером факультативного гетерохроматина является неактивная половая хромосома при гомогаметном кариотипе , например, неактивная X -хромосома у женских особей млекопитающих, деактивирующаяся в конденсированное гетерохроматиновое состояние; такая гетерохроматиновая X -хромосома наблюдается в интерфазе как тельце Барра . Вместе с тем, при гаметогенезе и на ранних стадиях эмбриогенеза обе X -хромосомы являются эухроматиновыми и транскрипционно активными.
Другим примером образования факультативного гетерохроматина является пахитенная стадия мейотического деления гетерогаметного гаметоцита, сопровождающаяся у млекопитающих при сперматогенезе образованием гетерохроматинового комплекса XY -хромосом — полового пузырька. Образование такого гетерохроматинового комплекса является временным и обратимым, необходимым для деактивации половых хромосом на этой стадии мейоза : в случае, когда X - и Y -хромосомы остаются активными на этой стадии, происходит нарушение баланса между продуктами экспрессии аутосом и половых хромосом, что приводит к гибели клеток.
Факультативный гетерохроматин обуславливает также «молчание» тканеспецифичных генов, переходящих в эухроматиновое состояние и экспрессирующихся только в дифференцированных клетках определённых тканей: в таких клетках активны ~10 % генов — остальные гены являются инактивированными и находятся в составе факультативного гетерохроматина.
Конститутивный (структурный) гетерохроматин содержится в обеих гомологичных хромосомах и локализован преимущественно в экспонированных участках — центромере , теломерах , ядрышковом организаторе . ДНК конститутивного гетерохроматина является преимущественно сателлитной ДНК , состоящей из тандемных повторов (например, HS1 (Human Satellite 1), HS2, HS3, альфа-сателлит и другие сателлиты человека). В интерфазном ядре конститутивный гетерохроматин образует хромоцентры с внутренней стороны ядерной мембраны, а также в районах ядрышковых организаторов. Вопрос о функциональной роли структурного гетерохроматина в эукариотической клетке остается открытым.
Хроматин является нуклеопротеидом — комплексом ДНК с гистонами . Конденсация хроматина в гетерохроматин сопровождается как модификацией гистонов, так и усложнением состава нуклеопротеидного комплекса за счёт участия в нём белков гетерохроматина HP1 (Heterochromatin Protein 1).
Гистоны гетерохроматинового комплекса характеризуются низкой степенью по лизиновым остаткам, что увеличивает их основные свойства и, соответственно, связывание с кислыми фосфатными группами ДНК, что способствует компактификации комплекса. Другой особенностью, ведущей к образованию гетерохроматина, является метилирование 27-го лизинового остатка гистона H3 белками Polycomb-комплекса 2 (PRC2) и 9-го лизинового остатка гистона H3 гистоновой метилтрансферазой Suv39h. Метилирование 9-го лизинового остатка гистона H3 ведёт к образованию сайта связывания гистона H3 и белка гетерохроматина HP1. У дрозофил метилтрансфераза Suv39h функционально ассоциирована с гистондеацетилазой таким образом, что ацетилированное и метилированное состояние 9-го лизинового остатка гистона H3 являются взаимоисключающими, то есть обеспечивается единый механизм деацетилирования и метилирования гистона H3, ведущий к усилению связывания с гистоном как ДНК, так и белка гетерохроматина HP1.
| Основное |

|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
|
||||||||||
|
Перестройки и
нарушения |
|||||||||||
|
Хромосомное
определение пола |
|||||||||||
| Методы | |||||||||||