Список земноводных России
- 1 year ago
- 0
- 0


Размножение земноводных — это способы размножения амфибий . Для большинства амфибий требуется вода: как для размножения, так и для жизни, хотя некоторые откладывают яйца на суше. Некоторые могут обитать в солоноватой воде (например, крабоядная лягушка ), но морских амфибий нет. Однако несколько сотен видов лягушек, обычно обитателей тёплых областей не нуждаются в воде для размножения в дикой природе. Они размножаются живорождением , экологической и эволюционной адаптации, которая позволила им стать полностью независимыми от стоячей воды. Почти все эти лягушки живут во влажных тропических лесах, и из их икринок вылупляются уже взрослые лягушата, проходя через стадию головастика внутри яйца. У большинства червяг и хвостатых земноводных оплодотворение, как правило, внутреннее. Бесхвостые амфибии , напротив, размножаются наружным оплодотворением, оставаясь в амплексусе .
Для размножения большинству земноводных требуется пресная вода, хотя некоторые откладывают яйца на суше. Некоторые могут обитать в солоноватой воде (например, крабоядная лягушка ), но морских амфибий нет. Однако несколько сотен видов лягушек, обычно обитателей тёплых областей, (например, Eleutherodactylus , Pacific Platymantis, живородящие жабы Nectophrynoides и многие другие тропические лягушки) не нуждаются в воде для размножения в дикой природе. Они размножаются живорождением, экологической и эволюционной адаптации, которая позволила им стать полностью независимыми от стоячей воды. Почти все эти лягушки живут во влажных тропических лесах, и из их икринок вылупляются уже взрослые лягушата, проходя через стадию головастика внутри яйца. Репродуктивный успех многих амфибий зависит не только от количества осадков, но и от времени года .
В тропиках многие земноводные размножаются непрерывно или в любое время года. В регионах с умеренным климатом размножение в основном сезонное, обычно весной, и вызвано увеличением продолжительности дня, повышением температуры или дождём. Эксперименты показали важность температуры при размножении амфибий .
Размножение безногих амфибий плохо изучено. У червяг оплодотворение внутреннее, самец выдавливает сперму в клоаку самки. Парные внутри мужской клоаки выделяют жидкость, напоминающую жидкость, вырабатываемую предстательной железой млекопитающих, которая может транспортировать и питать сперму. Оплодотворение, вероятно, происходит в яйцеводе .


Большинство саламандр также размножаются внутренним оплодотворением. Самец проталкивает сперматозоиды в клоаку самки . В большинстве случаев самец откладывает сперматофор , небольшую капсулу спермы поверх желеобразного конуса, на субстрат либо на суше, либо в воде. Самка берёт пакет со спермой, захватывая его губами клоаки и проталкивая в отверстие. Сперматозоиды перемещаются в сперматеку на клоаку, где остаются до овуляции , которая может наступить через много месяцев. Ритуалы ухаживания и методы передачи сперматофора различаются у разных видов. У некоторых сперматофор может быть помещён непосредственно в клоаку самки, в то время как у других самка может направляться к сперматофору или удерживаться объятиями, называемыми амплексусом. Некоторые примитивные саламандры из семейств Sirenidae , Hynobiidae и Cryptobranchidae размножаются внешним оплодотворением так же, как и лягушки: самка откладывает яйца в воду, а самец выделяет сперму на яйцеклетку .

За некоторыми исключениями, лягушки размножаются внешним оплодотворением. Как правило, самцы обычно прибывают первыми к месту размножения, призывают кваканьем самок. Другие самцы спокойно остаются поблизости, ожидая возможности отобрать самку. Самки выбирают партнёра и откладываются яйца. Самец крепко обхватывает самку передними конечностями либо за передними, либо перед задними лапами, или вокруг шеи. Они остаются в амплексусе , их клоаки расположены близко друг к другу. Самка откладывает яйца, а самец покрывает их спермой. Шероховатые подушечки на лапках самца помогают удерживать хватку. Часто самец собирает и удерживает яичную массу, образуя задними лапами подобие корзины. Хвостатая лягушка размножается внутренним оплодотворением. «Хвост» есть только у самца и используется для оплодотворения самки. Эта лягушка живёт в ручьях с быстрым течением, и внутреннее оплодотворение сохраняет сперму, не унося её водой . У бесхвостых наблюдаются два типа брачного поведения: сезонный и спонтанный. При первом типе, характерном для большинства видов бесхвостых, в определённые периоды года взрослые лягушки собираются для размножения в прудах, озёрах или ручьях. Многие бесхвостые возвращаются к тому водоёму, в котором они развивались головастиками. В результате можно наблюдать сезонные миграции , в которые вовлечены тысячи особей. При спонтанном типе размножения взрослые лягушки прибывают на места размножения, подталкиваемые некой внешней причиной (например, выпадением осадков в засушливой местности). В данном случае размножение и развитие икринок и головастиков происходит с максимальной скоростью, до того, как недолговечные водоёмы (лужи) успеют высохнуть .

Самцы видов бесхвостых с сезонным типом брачного поведения обычно первыми прибывают в места размножения и остаются там продолжительное время, призывая кваканьем самок и защищая свою территорию от других самцов. Самки появляются позже лишь на время спаривания и икрометания. В этой ситуации количество самцов у кромки воды всегда превышает количество самок, поэтому среди самцов существует жёсткая конкуренция. Крупные и сильные самцы издают более низкие звуки и захватывают самые привлекательные для размножения места. Наблюдения показали, что самки предпочитают именно таких самцов . У некоторых видов самцы могут применять технику перехвата. У них нет своей территории и они не квакают, а перехватывают самок, находящихся на пути к зовущему самцу. Поскольку кваканье требует больших затрат энергии, иногда роли меняются и зовущий самец оставляет свою территорию и становится перехватчиком .
У видов со спонтанным типом брачного поведения (например, рода лопатоногов Scaphiopus ) основной упор делается не на выбор партнёра, а на скорость размножения. Соответственно меняется и тактика. Самец, первым обнаруживший подходящее место (лужу), начинает громко призывать сородичей, на его зов сбегаются все окрестные представители обоих полов, которые тут же подхватывают призыв, создавая хор, слышный издалека. Иногда подходящие условия отсутствуют годами, и, соответственно, эти виды не размножаются несколько лет подряд. Самки вида обычно вымётывают за один брачный сезон около половины имеющейся у них в наличии икры. Предположительно, эта стратегия рассчитана на появление более подходящих условий для размножения в будущем .

За исключением некоторых представителей рода древолазов , образующих на время брачного сезона моногамные пары, все бесхвостые полигамны .
Как правило, процесс спаривания заключается в том, что самец осёдлывает самку в воде и обхватывает её туловище. Самка мечет икру, а самец покрывает её своей спермой (внешнее оплодотворение). У некоторых видов самцы могут задерживать икринки задними лапами на время, необходимое для оплодотворения. В этом плане уникальными среди бесхвостых являются виды западноафриканского рода Nimbaphrynoides , которые размножаются живорождением , и виды танзанийского рода Nectophrynoides , практикующие яйцеживорождение . В обоих случаях имеет место внутреннее оплодотворение, а самка рожает полностью развитых молодых лягушек (сеголеток), которые минуют стадию головастика .
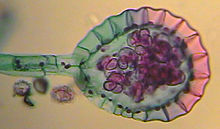
Лягушачья икра, как правило, запакована в многослойный желатиновый материал, предоставляющий икринкам некоторую защиту и не препятствующий прохождению кислорода , углекислого газа и аммиака . Эта защитная оболочка впитывает влагу и разбухает в воде. После оплодотворения внутренняя часть икринки разжижается, что обеспечивает развивающемуся зародышу свободу движений. У некоторых видов (например, красноногой лягушки и Rana sylvatica ) в желатиновом материале присутствуют одноклеточные зелёные водоросли . Было выдвинуто предположение, что они положительно влияют на развитие зародыша, повышая концентрацию кислорода, образующегося в процессе фотосинтеза . У большинства видов икринки чёрного или тёмно-коричневого цвета, что позволяет им нагреваться под лучами солнца сильнее окружающей среды. Так, например, температура внутри кластера икры Rana sylvatica была на 6 °C выше температуры воды, что способствовало быстрому развитию зародышей .

Большинство лягушек в период размножения можно классифицировать как активных и агрессивных. Как правило, лягушки собираются в месте размножения (небольшом озере или болоте), причём самцы обычно прибывают первыми, призывают самок, а также обустраивают территории. Другие самцы-спутники спокойно остаются поблизости, ожидая возможности захватить уже обустроенную территорию другого самца. Самки приплывают несколько позже мужских особей, происходит выбор партнёра и откладываются яйца. Самки уходят, и территории могут переходить от одного экземпляра к другому. Появляется больше самок, и со временем сезон размножения подходит к концу. С другой стороны, агрессивные лягушки встречаются там, где после дождя в засушливых регионах появляются временные лужи. Эти роющие лягушки обычно появляются после сильных дождей и собираются в месте размножения. Туда их привлекает призыв первого самца найти подходящее место, возможно, лужицу, которая образуется в одном и том же месте во время каждого сезона дождей. Собравшиеся лягушки могут кричать в унисон , и начинается бешеная активность, когда самцы пытаются спариться с обычно меньшим количеством самок .
У саламандр и тритонов существует прямая конкуренция между самцами за внимание самок, с тщательно продуманными демонстрациями ухаживания, чтобы удерживать внимание самки достаточно долго, чтобы она заинтересовалась выбором его для спаривания. Некоторые виды хранят сперму в течение долгих сезонов размножения .
Земноводные относятся к анамниям . Их яйца лишены зародышевых оболочек, в связи с чем для развития им требуются постоянное увлажнение .


Подавляющее большинство земноводных откладывают икру в пресных водоёмах, однако известны и исключения: червяги откладывают яйца в почву и обвивают место закладки, тем самым увлажняя яйца собственной слизью, , гигантские саламандры , и некоторые другие амфибии делают кладку на суше. Даже в этих случаях яйца нуждаются в повышенной влажности окружающей среды, обеспечение которой ложится на родителя. Известны виды, которые носят икринки на своём теле: самка прикрепляет их к животу, а самцы жаб-повитух обматывают шнурообразную кладку вокруг задних ног. Особенно необычно выглядит забота о потомстве у суринамской пипы — оплодотворённая икра вдавливается самцом в спину самки и последняя носит её на себе, пока из икры не вылупятся молодые пипы. В яйце зародыш подвешен в перивителлиновой жидкости и окружён полупроницаемыми студенистыми капсулами, а желточная масса обеспечивает питательные вещества. Когда личинки вылупляются, капсулы растворяются ферментами, выделяемыми железой на кончике рыла. Яйца некоторых саламандр и лягушек содержат одноклеточные зелёные водоросли. Они проникают в желейную оболочку после откладывания яиц и могут увеличить снабжение эмбриона кислородом посредством фотосинтеза. Похоже, что они ускоряют развитие личинок и снижают смертность. Яйца могут откладываться по одному, несколько или длинными нитями. Места для откладывания яиц: вода, грязь, норы, мусор и растения, а также некоторые амфибии откладывают икру под брёвнами или камнями .
Размер и форма агломерата икры характерна для каждого вида. Лягушки семейства Ranidae имеют тенденцию к шарообразным кластерам, а жабы к длинным цилиндрическим кластерам. Крошечный кубинский свистун откладывает икринки по одной и зарывает их во влажную почву. Leptodactylus pentadactylus создаёт пенное гнездо в норе, откладывая в него около тысячи икринок. Головастики появляются на свет, когда вода заполняет нору, а иногда развитие полностью происходит в гнезде. Красноглазая квакша откладывает икринки на листья, расположенные над поверхностью водоёма. Вылупляясь, головастики падают с листьев в воду. У некоторых видов на определённой стадии развития зародыши в икринках могут уловить вибрации, вызываемые хищниками (осами, змеями), и вылупиться раньше времени, чтобы приобрести мобильность и избежать гибели . В общем, длительность стадии развития зародышей в икре зависит от конкретного вида и окружающих условий. Как правило, головастики вылупливаются в течение недели после того, как капсула икринки распадается под воздействием гормона , вырабатываемого зародышем .

Из икринок вылупляются личинки (у бесхвостых земноводных называемые головастиками ), ведущие водный образ жизни. По своему строению личинки напоминают рыб: у них отсутствуют парные конечности, дышат жабрами (наружными, затем внутренними) и кожей; имеют двухкамерное сердце и один круг кровообращения, органы боковой линии . Некоторые червяги, альпийские саламандры и некоторые африканские живородящие жабы являются живородящими. Их личинки питаются железистым секретом и развиваются в яйцеводе самки, часто в течение длительного времени. Другие земноводные, но не червяги, яйцеживородящие. Яйца сохраняются в теле родителей или на нём, но личинки питаются желтками своих яиц и не получают питания от взрослых особей. Личинки появляются на разных стадиях своего роста, до или после метаморфоза, в зависимости от их вида .
Головастики, развивающиеся в прудах и ручьях, обычно являются травоядными. У прудовых головастиков, как правило, большие тела, большие хвостовые плавники и маленькие рты; они плавают в тихих водах, питаясь растительностью. У обитателей ручья более крупный рот, мелкое тело и хвостовые плавники; они прикрепляются к растениям и камням и питаются водорослями и мхами. У них относительно длинный спиралевидный кишечник, позволяющий им переваривать эту диету. Некоторые виды плотоядны на стадии головастика, питаются насекомыми, более мелкими головастиками и рыбой. Детёныши некоторых лягушек могут быть каннибалами: молодые головастики нападают на более крупного, более развитого головастика, когда тот превращается во взрослое животное .

Большинство видов червяг живородящи. Типичным из них является Typhlonectes Compressicauda , вид из Южной Америки. Одновременно в яйцеводе может развиваться до девяти личинок. Они довольно длинные, имеют парные мешкообразные жабры, маленькие глаза и специализированные зубы-скребки. Сначала они питаются желтками яиц, но по мере того, как этот источник питания иссякает, они начинают грызть реснитчатые эпителиальные клетки, выстилающие яйцеводы. Это стимулирует секрецию жидкости, богатой липидами и мукопротеинами, которыми они питаются вместе с соскобами со стенки яйцеводов. Они могут увеличить свою длину в шесть раз и быть в два раза длиннее своей матери до рождения. К этому времени они претерпели метаморфоз, потеряли глаза и жабры, у них развилась более толстая кожа и ротовые щупальца, а зубы снова исчезли и рассосались. Постоянный набор зубов вырастает вскоре после рождения .
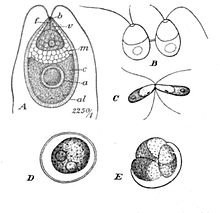
В отличие от других амфибий, головастики лягушек не похожи на взрослых особей . Личинки бесхвостых, появляющиеся из икринок, известны как «головастики». Они ведут полностью водный образ жизни, но известно одно исключение — головастики вида Nannophrys ceylonensis полусухопутные и обитают среди мокрых камней . Их тело, как правило, имеет овальную форму, хвост длинный, сплющенный по вертикали, приспособлен для плаванья. У головастиков хрящевой скелет, глаза лишены век, есть боковая линия , для дыхания служат жабры . В начале у головастиков появляются внешние жабры, а позже внутренние (жаберный мешок закрывает жабры и передние ноги). Развивающиеся лёгкие служат дополнительным дыхательным органом. Некоторые виды проходят метаморфоз ещё в икринке, и из икринок вылупляются лягушата. У головастиков нет настоящих зубов, но у многих видов челюсти покрыты параллельными рядами кератиновых зубчиков (два ряда на верхней челюсти, три ряда на нижней и роговой клюв). Количество рядов и точная морфология рта варьируют у разных видов и могут служить диагностическим признаком. Головастики семейства пиповые (за исключением рода Hymenochirus ) имеют пару передних усиков, делающих их похожими на небольших сомиков . Головастики имеют хрящевой скелет, жабры для дыхания (сначала внешние жабры, позже внутренние), систему боковых линий и большие хвосты, которые они используют для плавания .

После завершения стадии головастика бесхвостые проходят процесс метаморфоза , во время которого системы организма резко перестраиваются во взрослую форму. При метаморфозе происходят быстрые изменения в организме, так как полностью меняется образ жизни лягушки. Как правило, метаморфоз длится около суток. Он начинается с выработки гормона тироксина , который влияет на развитие тканей. Так, изменение органов дыхания включает в себя развитие лёгких параллельно с исчезновением жабр и жаберного мешка. Передние конечности становятся видимыми. Нижняя челюсть приобретает характерную для хищника форму, кишечник укорачивается. Нервная система адаптируется под стереоскопическое зрение и слух, а также под новые способы передвижения и питания. Глаза сдвигаются выше, формируются веки и связанные с ними железы. Модифицируются органы слуха (появляется слуховая перепонка и среднее ухо). Кожа становится толще и крепче, боковая линия исчезает (у большинства видов), формируются кожные железы. На последнем этапе метаморфоза исчезает хвост, ткани которого идут на развитие конечностей .Спиралевидный рот с роговыми зубными гребнями резорбируется вместе со спиральной кишкой. У животного развивается большая челюсть, и его жабры исчезают вместе с жаберным мешком. Быстро растут глаза и ноги, формируется язык. Есть связанные изменения в нейронных сетях, такие как развитие стереоскопического зрения и потеря системы боковой линии. Через несколько дней хвост реабсорбируется из-за более высокой концентрации тироксина, необходимой для этого .
У тритонов и саламандр метаморфозы менее интересны, чем у лягушек. Это связано с тем, что личинки уже плотоядны и продолжают питаться как хищники, когда становятся взрослыми, поэтому их пищеварительная система не изменяется. Их лёгкие начинают функционировать рано, работают они довольно плохо, но головастики не дышат ими так активно, как головастики лягушек. Другие метаморфозы, это уменьшение размера или потерю хвостовых плавников, закрытие жаберных щелей, утолщение кожи, развитие век и определённые изменения в зубном ряду и строении языка. Наибольшую уязвимость при трансформации проявляют хвостатые земноводные, так как развиваемая ими скорость плавания значительно падает, а длинный хвост, медленно проходящий различные метаморфозы является препятствием на суше . Трансформация может произойти примерно через 3 месяца . Жабры никогда не закрываются, они рассасываются непосредственно перед выходом из воды. Другие изменения включают уменьшение размера или потерю спинного плавника, закрытие жаберных щелей, утолщение и укрепление кожи, развитие век и некоторые изменения в зубном ряду и строении языка. Взрослые хвостатые весной и летом возвращаются в воду, а зимой живут на суше. Для адаптации к воде необходим гормон пролактин, а адаптация к наземным условиям зависит от тироксина . Наружные жабры, рассосавшиеся до первого выхода на сушу, после возвращения в воду не восстанавливаются .
При вылуплении типичная личинка саламандры имеет глаза без век, зубы как в верхней, так и в нижней челюсти, три пары перистых наружных жабр и длинный хвост со спинным и брюшным плавниками. Передние конечности могут быть частично развиты, а задние конечности могут быть рудиментарными у видов, живущих в пруду, но могут быть более развиты у видов, которые размножаются в движущейся воде. Личинки прудового типа часто имеют пару балансиров, стержнеобразных структур по обе стороны головы, которые могут предотвратить засорение жабр отложениями . Оба они способны размножаться . Саламандры без лёгких из семейства Plethodontidae ведут наземный образ жизни и откладывают небольшое количество непигментированных яиц скоплением среди влажной опавшей листвы. Каждое яйцо имеет большой желточный мешок, и личинка питается им, пока развивается внутри яйца, полностью сформировавшись в молодую саламандру. Самка саламандры часто высиживает яйца. Было замечено, что у представителей рода Ensatinas самка обвивается вокруг них и прижимается к ним областью горла, массируя их слизистым секретом .