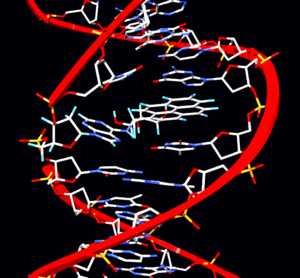
Точечная мутация
— тип мутации в
ДНК
или
РНК
, для которого характерна замена одного азотистого основания другим. Термин также применяется и в отношении парных замен, инсерции или делеции одного или нескольких
нуклеотидов
. Точечные мутации, возникающие в некодирующей ДНК, обычно никак себя не проявляют. Точечный мутант — организм, в генотипе которого произошла точечная мутация.
Точечные мутации классифицируют по эффекту, который изменённый нуклеотид оказывает на
триплет
:
Нонсенс-мутация
— мутация, в результате которой
кодон
теряет способность кодировать какую-либо аминокислоту и становится
стоп-кодоном
, что приводит к преждевременной терминации синтеза белка.
Миссенс-мутация
— переключает кодон на кодирование другой аминокислоты.
Содержание
Мутации замены оснований
Точечные мутации
замены оснований
классифицируют на транзиции и трансверсии
.
Трансверсия
— пуриновое основание замещается на пиримидиновое основание или наоборот.
Транзиции происходят чаще, чем трансверсии
.
Мутации замены оснований делятся на мишенные мутации замены оснований, немишенные и задерживающиеся мутации.
Мишенные мутации замены оснований
Мутации замены оснований, которые появляются напротив повреждений молекулы
ДНК
и способны останавливать синтез ДНК, называются мишенными мутациями замены оснований (от слова «мишень»)
. К мишенным мутациям замены оснований приводят, например, циклобутановые
пиримидиновые димеры
.
Немишенные мутации замены оснований
Иногда
мутации
замены оснований образуются на неповрежденных участках
ДНК
. Такие мутации называются немишенными мутациями замены оснований
. Механизмы образования немишенных мутаций замены оснований были разработаны в рамках полимеразной и полимеразно-таутомерной моделей мутагенеза.
Задерживающиеся мутации замены оснований
Мутации
замены оснований не всегда образуются сразу же после воздействия
мутагена
. Иногда они появляются после десятков циклов
репликации
. Это явление носит название задерживающихся
мутаций
.
Нестабильность генома
- главная причина образования
злокачественных опухолей
- характеризуется резким возрастанием количества
немишенных
и задерживающихся мутаций
. Механизмы образования задерживающихся мутаций в настоящее время не известны.
Делеция — в молекуле ДНК выпадает один или несколько
нуклеотидов
.
Инсерция — в молекулу ДНК встраивается один или несколько нуклеотидов.
Точечные мутации противопоставляются сложным мутациям, при которых один участок ДНК заменяется участком другой длины и иного нуклеотидного состава
.
Точечные мутации могут появляться напротив таких повреждений молекулы ДНК, которые способны останавливать синтез ДНК. Например, напротив циклобутановых пиримидиновых димеров. Такие мутации называются мишенными мутациями (от слова «мишень»)
. Циклобутановые пиримидиновые димеры вызывают как мишенные мутации замены оснований, так и мишенные мутации сдвига рамки
.
Иногда точечные мутации образуются на, так называемых, неповреждённых участках ДНК, часто в небольшой окрестности от фотодимеров. Такие мутации называются немишенными мутациями замены оснований или немишенными мутациями сдвига рамки
.
Точечные мутации образуются не всегда сразу же после воздействия мутагена. Иногда они появляются после десятков циклов репликации. Это явление носит название задерживающихся мутаций
. При нестабильности генома, главной причине образования злокачественных опухолей, резко возрастает количество немишенных и задерживающихся мутаций
.
Причины возникновения
Точечные мутации могут возникать в результате спонтанных мутаций, происходящих во время
репликации ДНК
. Они также могут возникать в результате действия
мутагенов
— к примеру, воздействия
ультрафиолетового
или
рентгеновского излучения
, высокой температуры или химических веществ. Мутации появляются при синтезе молекулы ДНК, содержащей повреждения, в процессах репликации ДНК, репарации ДНК или транскрипции
.
В настоящее время существует несколько подходов, используемых для объяснения природы и механизмов образования точечных мутаций. В рамках общепринятой, полимеразной модели считается, что единственная причина образования мутаций замены оснований — спорадические ошибки ДНК-полимераз
. Уотсон и Крик
предложили таутомерную модель спонтанного мутагенеза. Они объяснили появление спонтанных мутаций замены оснований тем, что при соприкосновении молекулы ДНК с молекулами воды могут изменяться таутомерные состояния оснований ДНК. Образование мутаций замены оснований объяснялось образованием Хугстиновских пар
. Одной из причин образования мутаций замены основания считается дезаминирование 5-метилцитозина
.
Примечания
Тарасов В. А. Молекулярные механизмы репарации и мутагенеза. — М.: Наука, 1982. — 226 с.
Friedberg E. C., Walker G. C., Siede W. DNA repair and mutagenesis. — Washington: ASM Press, DC, 1995.
Fields Virology by David M Knipe and Peter M Howley | Publisher: Lippincott Williams & Wilkins, 2007 |
ISBN 0-7817-1832-5
| стр. 394
от 22 октября 2018 на
Wayback Machine
Transition versus Transversion mutations
↑
Wang C.-I., Taylor J.-S. In vitro evidence that UV-induced frameshift and substitution mutations at T tracts are the result of misalignment-mediated replication past a specific thymine dimer // Biochemistry - 1992. – 31. – P. 3671–3681.
↑
Lawrence C. W., Banerjee S. K., Borden A., LeClerc J. E. T-T cyclobutane dimers are misinstructive rather than non-instructive, mutagenic lesions // Mol. Gen. Genet. – 1990. – 222. – P. 166-169.
↑
LeClerc J. E., Borden A., Lawrence C. W. The thymine-thymine pyrimidine-pyrimidine (6-4) ultraviolet light photoproduct is highly mutagenic and specifically induces 3'-thymine-to-cytosine transitions in Escherichia coli // Proc. Nat. Acad. Sci. USA. – 1991. – 88. – P. 9685-9686.
Maor-Shoshani A., Reuven N. B., Tomer G., Livneh Z. Highly mutagenic replication by DNA polymerase V (UmuC) provides a mechanistic basis for SOS untargeted mutagenesis // Proc. Natl. Acad. Sci. USA - 2000. - 97. – P. 565–570.
Little J. B., Gorgojo L., Vetrovs H. Delayed appearance of lethal and specific gene mutations in irradiated mammalian cells // Int. J. Radiat. Oncol. Biol. Phys. — 1990. — 19. — P. 1425—1429.
Кордюм В. И. Опухоль – как она видится сегодня с позиций молекулярной генетики // Биополимеры и клетка. – 2001. – Т. 17. – С. 109-139.
Niwa O. Radiation induced dynamic mutations and transgenerational effects // J. Radiation Research. — 2006. — 47. — P. B25-B30.
Ауэрбах Ш. Проблемы мутагенеза. — М.: Мир, 1978. — 463 с.
Friedberg E. C., Walker G. C., Siede W., Wood R. D., Schultz R. A., Ellenberger T. DNA repair and mutagenesis. — part 3. Washington: ASM Press. — 2006. 2nd ed.
Levine J. G., Schaaper R. M., De Marini D. M. Complex frameshift mutations mediated by plasmid pkm 101: Mutational mechanisms deduced mutation spectra in Salmonella // Genetics. — 1994. — 136. — P. 731—746.
Pham P., Bertram J. G, O’Donnell M., Woodgate R., Goodman M. F. A model for SOS-lesion-targeted mutations in Escherichia coli // Nature. — 2001. — 408. — P. 366—370.
Wang C.-I., Taylor J.-S. In vitro evidence that UV-induced frameshift and substitution mutations at T tracts are the result of misalignment-mediated replication past a specific thymine dimer // Biochemistry — 1992. — 31. — P. 3671-3681.
Maor-Shoshani A., Reuven N. B., Tomer G., Livneh Z. Highly mutagenic replication by DNA polymerase V (UmuC) provides a mechanistic basis for SOS untargeted mutagenesis // Proc. Natl. Acad. Sci. USA — 2000. — 97. — P. 565—570.
Little J. B., Gorgojo L., Vetrovs H. Delayed appearance of lethal and specific gene mutations in irradiated mammalian cells // Int. J. Radiat. Oncol. Biol. Phys. — 1990. — 19. — P. 1425—1429.
Niwa O. Radiation induced dynamic mutations and transgenerational effects // J. Radiation Research. — 2006. — 47. — P. B25-B30.
Banerjee S. K., Borden A., Christensen R. B., LeClerc J. E., Lawrence C. W. SOS-dependent replication past a single trans-syn T-T cyclobutane dimer gives a different mutation spectrum and increased error rate compared with replication past this lesion in uniduced cell // J. Bacteriol. — 1990. — 172. — P. 2105—2112.
Jonczyk P., Fijalkowska I., Ciesla Z. Overproduction of the subunit of DNA polymerase III counteracts the SOS-mutagenic response of Esthetician coli // Proc. Nat. Acad. Sci. USA. — 1988. — 85. — Р. 2124—2127.
Tang M., Shen X., Frank E. G., O’Donnell M., Woodgate R., Goodman M. F. UmuD’(2)C is an error-prone DNA polymerase. Escherichia coli pol V // Proc. Natl. Acad. Sci. USA — 1999. — Vol. 96. — P. 8919-8924.
Watson J. D., Crick F. H. C. The structure of DNA // Cold Spring Harbor Symp. Quant. Biol. — 1953. — 18. — P. 123—131.
Полтев В. И., Шулюпина Н. В., Брусков В. И. Молекулярные механизмы правильности биосинтеза нуклеиновых кислот. Компьютерное изучение роли полимераз в образовании неправильных пар модифицированными основаниями // Молек. биол. — 1996. — 30. — С. 1284—1298.
Cannistraro V. J., Taylor J. S. Acceleration of 5-methylcytosine deamination in cyclobutane dimers by G and its implications for UV-induced C-to-T mutation hotspots // J. Mol. Biol. — 2009. — 392. — P. 1145—1157.
Ссылки
Марри Р., Греннер Д., Мейес П., Родуэлл В.
Биохимия человека. —
М.
: «Мир», 1993. — Т. 2. — С. 98. — 415 с. —
12 000 экз.
—
ISBN 5-03-001775-5
.