Interested Article - Дигенетические сосальщики

- 2021-02-12
- 1

Дигенетические сосальщики ( лат. Digenea ) — подкласс паразитических плоских червей из класса трематод (Trematoda) . Их жизненный цикл , за редкими исключениями, протекает в нескольких хозяевах и сопровождается закономерным чередованием не менее трёх поколений. Описано около 7200 видов. Некоторые трематоды (около 40 видов) — паразиты человека, вызывающие опасные заболевания — трематодозы , например, шистосомоз и описторхоз . К трематодам относятся печёночная двуустка ( Fasciola hepatica ), кошачья двуустка ( Opisthorchis felineus ), шистосомы ( Schistosoma ), Leucochloridium paradoxum .
Раньше, особенно в русскоязычной научной традиции, название трематоды (Trematoda) использовалось как синоним дигенетических сосальщиков . В современной гельминтологии этим термином называют класс плоских червей, объединяющий подклассы аспидогастрей (Aspidogastrea) и дигенетических сосальщиков (Digenea) .
Описание
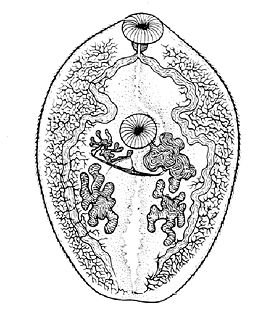
К подклассу дигенетических сосальщиков относятся самые мелкие представители плоских червей, взрослая особь мелких видов может достигать 0,2 мм, крупных видов — 6 см .

Жизненный цикл
Дигенетические сосальщики обладают сложным жизненным циклом с гетерогонией : закономерным чередованием амфимиктной (обычно гермафродитной ) и партеногенетических стадий. Всех представителей класса характеризует наличие стадий, паразитирующих в беспозвоночных (обычно в брюхоногих моллюсках ). У большинства видов в жизненном цикле также присутствует, по меньшей мере, ещё один хозяин — позвоночное .

Партеногенетические поколения
Из яйца выходит особь первого партеногенетического поколения — мирацидий , который способен плавать с помощью биения ресничек . Для дальнейшего развития мирацидий должен заразить следующего хозяина, в котором будут паразитировать все партеногенетические стадии. В роли этого хозяина практически у всех дигенетических сосальщиков выступают брюхоногие моллюски. Лишь некоторые группы в качестве хозяина используют двустворчатых моллюсков (например, из семейства ) или многощетинковых червей (из семейства ). Эту стадию характеризует высокая специфичность к хозяину: как правило, мирацидии заражают и развиваются в моллюсках лишь одного вида (или нескольких близкородственных видах).
Выход мирацидия из яйца у разных видов происходит либо после заглатывания яйца хозяином ( пассивное заражение ), либо во внешней среде ( активное заражение ; подразумевает поиск хозяина). Последний вариант может реализоваться только у тех видов, яйца которых для развития должны попасть в воду. В то же время, заражение даже водных моллюсков некоторыми видами трематод осуществляется пассивно.
При обнаружении хозяина мирацидий проникает через покровы и претерпевает метаморфоз , обычно сводящийся к сбрасыванию ресничных покровов и формированию нового эпителия — тегумента . У небольшого числа видов превращение носит более кардинальный характер, и их мирацидии теряют почти все органы, а следующие стадии развиваются из небольшого числа клеток. Претерпевший метаморфоз мирацидий называют материнской спороцистой . В материнской спороцисте из партеногенетических яйцеклеток развивается следующее поколение — дочерние партениты , в зависимости от строения называемые редиями или дочерними спороцистами . Дочерние партениты в размножения отрождают либо следующие поколения дочерних партенит такого же строения, либо церкарий — личинок амфимиктного поколения.
Дочерние партениты некоторых трематод могут обитать в моллюске в течение нескольких лет, не приводя к его гибели, подавляя, однако, его репродуктивную активность. Паразитизм других видов, характеризующихся массовым выходом церкарий, закономерно приводит к гибели хозяина.
Амфимиктное поколение
Поведение вышедшей из хозяина церкарии (расселительной личинки) специфично для разных видов. Выделяют три основных стратегии:
- прямое заражение окончательного хозяина через наружные покровы или при случайном заглатывании;
- проникновение и инцистирование в промежуточном хозяине (стадия метацеркарии ), при этом промежуточный хозяин служит пищей для окончательного хозяина или следующего промежуточного хозяина (встраивание в трофическую цепь );
- инцистирование во внешней среде на неживых предметах, водных растениях или раковинах моллюсков (стадия адолескарии ) с последующим пассивным заражением окончательного хозяина.
Существуют дигенетические сосальщики (например, лейкохлоридий парадоксальный ), церкарии которых не выходят из тела хозяина, а заражают следующего хозяина при поедании им предыдущего.
За исключением единичных случаев, окончательный хозяин, в котором развиваются половозрелые особи амфимиктного поколения ( мариты ), — позвоночное. В организме хозяина они могут локализоваться в кишечнике , паренхиме и протоках печени , жёлчном пузыре , органах выделительной системы , дыхательных путях, кровеносном русле или даже под веком . Образующиеся после скрещивания марит яйца выводятся во внешнюю среду с калом или мочой хозяина.
Гипотеза о социальной организации
В 2010 году американские исследователи показали, что партениты одного из видов рода , паразитирующие в улитках Cerithidea californica , чётко разделяются на две группы, характеризующиеся различными размерами, строением и поведением. Более крупные редии ведут малоподвижный образ жизни и способны к размножению, более многочисленные мелкие редии, названные первооткрывателями «солдатами», не размножаются и активно атакуют как плоских червей других видов, так и неродственных им представителей того же вида .
Однако, данная концепция подвергается критике со стороны многих специалистов по гельминтам. В качестве основных аргументов, демонстрирующих несостоятельность вывода о наличии у редий разделения на настоящие касты, можно привести следующие наблюдения:
- Между размерами тела особей из разных «каст» нет выраженного пробела, и они значительно перекрываются .
- У «редий-солдат» также присутствует зародышевый материал и потенциал к размножению .
Примечания
- ↑ Подкласс (англ.) в Мировом реестре морских видов ( World Register of Marine Species ). (Дата обращения: 30 октября 2018) .
- Дигенетические сосальщики // Дебитор — Евкалипт. — М. : Советская энциклопедия, 1972. — ( Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 8).
- / Чесунов А. В. // Телевизионная башня — Улан-Батор. — М. : Большая российская энциклопедия, 2016. — С. 364—365. — ( Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 32). — ISBN 978-5-85270-369-9 .
- Класс (англ.) в Мировом реестре морских видов ( World Register of Marine Species ). (Дата обращения: 30 октября 2018) .
- , p. 454.
- Hechinger R. F., Wood A. C., Kuris A. M. Social organization in a flatworm: trematode parasites form soldier and reproductive castes (англ.) // Proceedings of the Royal Society B. — 2011. — Vol. 278 . — P. 656—665 . — doi : .
- Николаев К. Е. Особенности реализации жизненных циклов трематод семейств Echinostomatidae и Renicolidae в литоральных экосистемах Кандалакшского залива Белого моря : Дис. канд. биол. наук. — СПб. : Зоологический институт РАН, 2012. — 300 с.
- Zikmundová, Jana. : [ англ. ] : Бакалаврская диссертация / Dr. Aneta Kostadinova, Ph.D.. — Czech Republic, České Budějovice : Faculty of Science, University of South Bohemia in České Budějovice, 2011. — 36 с.
Литература
- Белопольская М. М. Трематоды семейства Microphallidae Travassos, 1920 // Трематоды животных и человека. Основы трематодологии. Том 21. — М. : Издательство АН СССР , 1963. — С. 260—299. — 504 с. — 1500 экз.
- Гинецинская Т. А. Трематоды, их жизненные циклы, биология и эволюция / АН СССР . — Л. : Наука , Ленингр. отд-ние, 1968. — 412, [10] с.
- Галактионов К. В. , Добровольский А. А. / Рос. акад. наук, Зоол. ин-т, Кол. науч. центр РАН, Мурм. мор.-биол. ин-т. — СПб. : Наука , 1998. — 404, [16] с. — ISBN 5-02-026089-4 . (недоступная ссылка)
- Морозова Д. А. / Морозова Дарья Андреевна; (Место защиты: Ин-т проблем экологии и эволюции им. А. Н. Северцова РАН). — Борок, 2011. — 176 с. .
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Протисты и низшие многоклеточные // Зоология беспозвоночных: функциональные и эволюционные аспекты. — М. : Издательский центр «Академия», 2008. — Т. 1. — С. 449—460.
- , Ястребова И. В. . — М. : Товарищество научных изданий КМК, 2014. — 344 с. — ISBN 978-5-87317-971-8 .
|
|
|
|---|---|
| Таксономия |
|
| В библиографических каталогах |
|
|
Стадии жизненного цикла
(Digenea)
|
|
|---|---|
|
Яйцо
→
Мирацидий
→
Материнская спороциста
→
Редия
/
→
Церкария
→
Метацеркария
/
Адолескария
→
Марита
|
|
|
Примечание
: в жизненном цикле многих дигенетических сосальщиков отсутствуют стадии метацеркарии и адолескарии.
|
|
- 2021-02-12
- 1