, а чем больше значение
, а чем больше значение
 , тем меньше значение
, тем меньше значение
 (см. подробнее
(см. подробнее
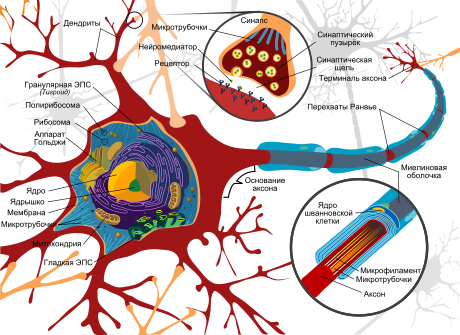
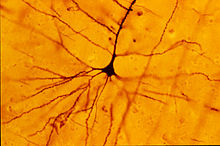
Нейрон
- 1 year ago
- 0
- 0

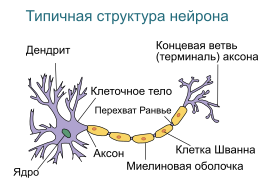

Дендрит (от греч. δένδρον (dendron) — дерево) — разветвлённый отросток нейрона , который получает информацию через химические (или электрические ) синапсы от аксонов (или дендритов и сомы ) других нейронов и передаёт её через электрический сигнал телу нейрона ( перикариону ), из которого вырастает. Термин «дендрит» ввёл в научный оборот швейцарский ученый В. Гис в 1889 году .
От сложности и разветвлённости дендритного дерева зависит то, сколько входных импульсов может получить нейрон. Поэтому одно из главных назначений дендритов заключается в увеличении поверхности для синапсов (увеличении рецептивного поля ), что позволяет им интегрировать большое количество информации, которая поступает к нейрону.

Огромное многообразие дендритных форм и разветвлений, как и открытые недавно различные виды дендритных нейромедиаторных рецепторов и потенциалзависимых ионных каналов (активных проводников), является свидетельством богатого разнообразия вычислительных и биологических функций, которые дендрит может выполнять в ходе обработки синаптической информации по всему мозгу.
С накоплением новых эмпирических данных становится все более очевидным, что дендриты играют ключевую роль в интеграции и обработке информации, а также способны генерировать потенциалы действия и влиять на возникновение потенциалов действия в аксонах , представая как пластичные , активные механизмы со сложными вычислительными свойствами. Исследование обработки дендритами синаптических импульсов является необходимым для понимания роли нейрона в обработке информации в ЦНС , а также и для выявления причин многих психоневрологических заболеваний.
Тело нейрона (сома) и дендриты — два главных участка нейрона, которые воспринимают входные импульсы от других нейронов. Согласно классической «нейронной доктрине», предложенной Рамоном-и-Кахалем , информация через большинство нейронов протекает в одном направлении (ортодромический импульс) — от дендритных ветвей и тела нейрона (которые являются рецептивными частями нейрона, к которым импульс входит) к единому аксону (который является эффекторной частью нейрона, с которой импульс начинается). Таким образом, большинство нейронов имеет два типа отростков (нейритов): один или более дендритов, реагирующих на входящие импульсы, и аксон, который проводит выходной импульс. Тем не менее, с открытием антидромических импульсов (которые протекают от аксона к соме и к дендритам) и дендритных потенциалзависимых каналов, представление о природе дендритов начало меняться.
Существенное различие между дендритными деревьями и другими подобными древовидными структурами в природе заключается в том, что дендритные имеют больше степеней свободы . Поэтому для того, чтобы понять влияние морфологии дендритов на их электрически-вычислительные свойства, следует принимать во внимание тот факт, что нейрон — это трёхмерная структура. Вследствие такой сложности роль дендритной структуры выходит далеко за пределы роли просто быть местом синаптических контактов . Уже для Рамона-и-Кахаля было очевидным, что разветвлённость дендритов отражает количество контактов, которые один нейрон может иметь с другими нейронами. Нейрон без дендритов, имея сферическое клеточное тело, обладал бы очень ограниченной площадью поверхности для получения входных импульсов. Поэтому дендриты являются как бы расширением тела нейрона, увеличивая его площадь поверхности без чрезмерного увеличения объёма клетки. Так, 97 % площади поверхности мотонейрона (исключая аксон) является дендритной.
Например, дендриты типичного моторного нейрона кошки занимают 370 000 мкм² площади поверхности, вместе с тем предоставляя 300 000 мкм² для синаптических входов . Чтобы предоставить эквивалентную поверхность, сферическое клеточное тело должно иметь диаметр 340 мкм, а объём - 20 000 000 мкм³ .
Тот факт, что 80 % площади поверхности ближайших к соме дендритов мотонейрона покрыто синапсами, свидетельствует, что увеличение площади поверхности действительно является значимым для увеличения количества входных импульсов из нейрона, вместе с тем позволяя вместить большее количество нейронов в непосредственной близости друг к другу и расширить их возможности для большего разнообразия аксонов от других нейронов .
| Аксоны | Дендриты |
|---|---|
| За редкими исключениями, каждый нейрон имеет аксоны. | Большинство нейронов имеет дендриты. |
| Аксоны появляются первыми во время нейрональной дифференциации. | Только когда аксоны сформировались, начинают дифференцироваться дендриты. |
| Начальные сегменты аксонов отделены специальной плазматической мембраной , обладающей высокой плотностью ионных каналов и особой организацией цитоскелета . | Дендриты неразделимы с соматической цитоплазмой, и точка перехода между ними трудноразличима. |
| Аксоны, как правило, имеют цилиндрическую форму с круглым или эллиптическим поперечным сечением. | Дендриты имеют конусообразную форму и небольшие отростки, которые придают ему неправильное поперечное сечение. |
| Большие аксоны имеют миелиновую оболочку, толщина которой пропорциональна размеру аксона. | Дендриты не миелированы, хотя редко некоторые из них имеют несколько слоев миелина. |
| Размер аксона зависит от нейрофиламента и количества микротрубочек с перевесом нейрофиламентов в крупных аксонах. | Дендритный цитоскелет является менее организованным, а микротрубочки преобладают даже в крупных дендритах. |
| Микротрубочки в аксонах имеют однородную полярность, с плюс-концами, удаленными от сомы. | Микротрубочки в ближайших дендритах имеют смешанную полярность, с плюс- и минус-концами, наружу направленными к соме. |
| Микротрубочки в аксонах обогащены тау-белками . | Дендриты могут иметь несколько тау-белков, но они имеют много протеинов , которых нет у аксонов. |
| Зрелые аксоны не имеют рибосом , хотя некоторое их количество можно обнаружить в начальном сегменте. | У дендритов есть эндоплазматический ретикулум и цитоплазматические полисомы, вместе с большим количеством специфических мРНК . |
| Ветви аксонов удалены от сомы | Дендриты начинают своё разветвление возле сомы |
| Ветви аксонов образуют тупые углы и имеют диаметры, схожие с родительским стволом. | Ветви дендритов образуют острые углы и имеют диаметры, меньшие родительского ствола. |
| Аксоны имеют пресинаптические образования, находящиеся на конце аксонных ветвей. | Дендриты имеют большое количество постсинаптических образований, которые выходят из дендритного ствола. |
| Потенциалы действия обычно возникают в аксонном холмике и активно распространяются от сомы. | Хотя дендриты способны генерировать потенциалы действия , чаще они модулируют электрическое состояние сомы и начального сегмента аксона. |
| Традиционно аксоны специализированы для проведения возбуждения и синаптической передачи сигнала, — нейронный «выход». | Дендритная архитектура лучше всего приспособлена для интеграции синаптических импульсов из множества входов, — нейронный «вход». |


В отличие от аксонов, дендриты имеют высокое содержание рибосом и образуют относительно локальные соединения, которые непрерывно ветвятся во все стороны и сужаются, что ведёт к уменьшению в размерах дочерних отростков на каждой ветке. Также, в отличие от ровной поверхности аксонов, поверхность большинства дендритов усеяна выступающими маленькими органеллами , которые называются дендритными шипиками и которые обладают высокой пластичностью: они способны рождаться и умирать, менять свою форму, объём и количество на протяжении короткого отрезка времени. Среди дендритов есть как те, которые усеяны шипиками ( пирамидальные нейроны ), так и те, которые шипиков не имеют (большинство интернейронов ), достигая максимального количества транзакций в клетках Пуркинье — 100 000 транзакций, то есть около 10 шипиков на 1 пм. Другая отличительная черта дендритов в том, что они характеризуются разным количеством контактов (вплоть до 150 000 на дендритном дереве в клетке Пуркинье) и различными типами контактов (аксон-шипик, аксон-ствол, дендро-дендритные).
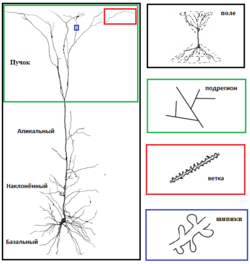
Одна из принятых, но и самых тяжёлых, в нейробиологии классификаций нейронов (основана Кахалем) основывается на их дендритной структуре и плотности. Однако, с другой стороны, есть нейроны, которые не имеют дендритов («адендритные нейроны»), а только разветвлённые аксоны (напр., нейроны дорсального корешкового ганглия и симпатического ганглия); есть нейроны со слишком сложными дендритами. К нейронным типам с наиболее характерными дендритными формами относятся :
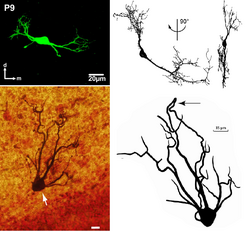
В связи с большим количеством типов нейронов и дендритов целесообразно рассмотреть морфологию дендритов на примере одного конкретного нейрона — пирамидальной клетки. Пирамидальные нейроны находятся во многих регионах мозга млекопитающих: гиппокамп , миндалина , неокортекс . Эти нейроны наиболее обильно представлены в коре головного мозга , составляя более чем 70—80 % всех нейронов изокортекса млекопитающих . Наиболее популярными, и поэтому лучше исследованными, являются пирамидальные нейроны 5 слоя коры: они получают очень мощный поток информации, которая прошла через различные предыдущие слои коры, и имеют сложную структуру на поверхности мягкой мозговой оболочки («апикальный пучок»), которая получает входные импульсы из иерархически обособленных структур; далее эти нейроны посылают информацию другим корковым и подкорковым структурам. Хотя, подобно другим нейронам, пирамидальные клетки имеют апикальные и базальные дендритные пучки, им также присущи дополнительные отростки вдоль апикальной дендритной оси — это т. н. «наклонённый дендрит» (), который раз или дважды ветвится от основания. Особенностью дендритов пирамидальных нейронов является также то, что они могут отправлять ретроградные сигнальные молекулы (напр., эндоканабиноиды ), которые проходят в обратном направлении через химический синапс к аксону пресинаптического нейрона .
Хотя часто дендритные ветви пирамидальных нейронов сравнивают с ветвями обычного дерева, они таковыми не являются. В то время как диаметр ветвей дерева постепенно сужается с каждым делением и становится все более коротким, диаметр последней ветви дендрита пирамидальных нейронов значительно тоньше, чем её родительская ветка, и эта последняя ветвь часто является самым длинным сегментом дендритного дерева. Тем более, диаметр оконечности дендрита не является суженным, в отличие от апикального ствола веток дерева: он имеет <1 мкм в диаметре, как в точке соединения с главным апикальным стволом, так и на своем удалённом конце, 100 мкм от ствола. Такая уникальная геометрия дендритов хорошо подходит для электрической сегментации и раздельной обработки информации в этих сегментах, поскольку здесь синаптические токи собраны с относительно большой площади в пределах региона с высоким сопротивлением. Поэтому даже относительно небольшой синаптический ток, поступающий в дендритные окончания, генерирует здесь значительную деполяризию, достаточную для порождения локального дендритного потенциала действия .

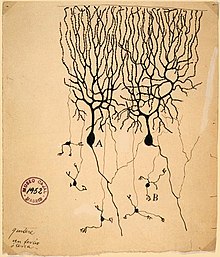
Своей функциональностью и высокой рецептивностью дендриты обязаны сложной геометрической разветвленности. Дендриты одного нейрона, взятые вместе, называются «дендритным деревом», каждая ветвь которого называется «дендритной ветвью». Хотя иногда площадь поверхности дендритной ветки может быть достаточно обширной, чаще всего дендриты находятся в относительной близости от тела нейрона (сомы), из которого выходят, достигая в длину не более 1—2 мкм. Количество входных импульсов, которые данный нейрон получает, зависит от его дендритного дерева: нейроны, которые не имеют дендритов, контактируют только с одним или несколькими нейронами, тогда как нейроны с большим количеством разветвлённых деревьев способны принимать информацию от множества других нейронов. Наиболее сложную и красивую дендритную форму имеют клетки Пуркинье , дерево на которых имеет около 400 верхушек, тогда как альфа-мотонейроны в спинном мозгу кошки имеют лишь 8—12 дендритных деревьев, каждое с около 30 верхушками .
Рамон-и-Кахаль , изучая дендритные разветвления, пришел к выводу, что филогенетические различия в специфических нейрональных морфологиях поддерживают отношения между дендритной сложностью и количеством контактов . Сложность и разветвлённость многих типов нейронов позвоночных (например, пирамидальные нейроны коры, клетки Пуркинье мозжечка , митральные клетки обонятельных луковиц ) растёт с увеличением сложности нервной системы . Эти изменения связаны как с необходимостью для нейронов формировать больше контактов, так и с необходимостью контактировать с дополнительными нейронными типами в конкретном месте нейронной системы.
Следовательно, способ связанности между нейронами является одним из наиболее фундаментальных свойств их разносторонних морфологий и именно поэтому дендриты, которые формируют одно из звеньев этих связей, определяют многообразие функций и сложность конкретного нейрона.
Впрочем, морфология дендритов определяет не только количество и тип контактов, которые может образовать данный нейрон с другими нейронами, но и его вычислительные операции. Как моделирование, так и электрофизиологические исследования поведения нейронов in vitro и in vivo , показали, что нейроны с более сложной морфологией (большие и разветвлённые дендритные дерева, больше веток и шипиков ) характеризуются пачечной пульсацией, тогда как нейроны с более простой морфологией имеют тенденцию к тоническим спайкам . Также морфология дендритов имеет существенное влияние на распространение потенциала действия , выявление дендритами совпадений во входных синаптических импульсах .

Трёхмерное распределение дендритных ветвей также имеет важное значение для определения типа информации, которая поступает к нейрону. Тогда как нейроны с дендритными деревьями, локализованными только в данном слое коры, могут получить очень ограниченное количество и разнообразие контактов, то нейроны с расширенными дендритными ветвями, простирающимися в другие слои (напр., как большие пирамидальные нейроны) способны получать значительно более разнообразные и сложные входные импульсы .
Но разветвленность и многообразие дендритной морфологии также имеет и ограничительное воздействие на нейронные высчисления, в частности на эффективность входных синаптических импульсов (синаптическую информационную эффективность, SIE ): нейроны с более развитыми дендритными деревьями характеризуются более резким и быстрым уменьшением синаптической эффективности (амплитуды синаптических потенциалов) вдоль пути от удалённого места синаптического входа до сомы, что приводит к затуханию постсинаптического потенциала . Впрочем, значительная нелинейность дендритов, которая обусловлена наличием потенциалзависимых ионных каналов , способна преодолеть это затухание, поддерживая и усиливая импульс на разных уровнях дендритного дерева .
Диаметр дендритов возле сомы имеет несколько мкм, становясь меньше 1 мкм, когда они постепенно ветвятся. Много типов дендритов усеяны множеством крошечных отростков, дендритными шипиками , которые образуют чрезвычайно тонкие (около 0,1 мкм) и короткие (1 мкм) дендритные веточки. Эти шипики является главной мишенью для возбудимых синаптических входов и играют важную роль в нейронной пластичности. Длина самих дендритных деревьев колеблется от очень коротких (100—200 мкм, как в шипиковых звёздчатых клетках коры млекопитающих) до достаточно больших (1—2 мм, как у спинного альфа-мотонейрона ). Общая длина дендритов может достигать 10 4 мкм (1 см) и более .
В целом, дендриты (не включая шипики) занимают примерно такой же объём коры , как и аксоны (без включения тела клеток и кровеносных сосудов ) (на примере коры мыши) :
В некоторых регионах ЦНС соседние дендритные деревья тесно переплетаются, очень часто деля между собой ту же область и группу аксонов, с которыми формируют синаптические контакты. Это особенно характерно для дендритов пирамидальных нейронов в коре, тогда как в других регионах мозга дендритные деревья (напр., клеток Пуркинье в мозжечке) отделены друг от друга и не перекрываются, каждый имея свои индивидуальные синаптические контакты. Именно поэтому на примере пирамидальных клеток лучше всего заметно значительное влияние морфологии дендритов на сложность нейронных операций.
Если предположить, что дендриты имеют цилиндрическую форму и средний диаметр 0,9 мкм (на основе данных электронной микрофотографии ), то длина дендритов в 1 мм³ составит 456 м, что почти на порядок меньше, чем длина аксонов в том же объёме, иллюстрируя большую среднюю толщину дендритов. Следовательно, 9,2×10 4 нейронов в 1 мм³ соответствовало бы ~276—460 м дендритам в 1 мм³ .
Из этого следует, что в системе связей между пирамидальными нейронами, которая включает большинство синапсов коры, влияние одного нейрона на другой является достаточно слабым, опосредуясь в основном одним синапсом. Это означает, что сколько каждый кортикальный пирамидальный нейрон имеет синапсов (~4000), со столькими же другими пирамидальными нейронами он образует контакты .
Подобному максимальному расхождению сигналов от одной клетки к тысячам других соответствует такое же максимальное восхождение: тысячи синапсов на дендритном дереве одной пирамидальной клетки (при 3,3 синапсах на мкм дендритной длины) формируют контакты с аксонами стольких же различных пирамидальных клеток .
Такая уникальная дендритная морфология пирамидальных клеток, которая позволяет им формировать не похожую на другие нейроны сеть контактов, привела некоторых исследователей к мысли, что «кора предстаёт как „устройство“ для наиболее широкого распространения и наиболее глубинного смешивания сигналов, совместного с естественными ограничениями нервных клеток» .
Морфология дендритов придаёт им уникальные электрические и пластические свойства, и, как следствие, обусловливает вариации в вычислительных свойствах самих нейронов. Если, например, рассматривать дендриты как пассивные проводники (классическое представление), то разнообразная дендритная морфология влияет только на частоту нейронного разряда ( англ. firing). Однако, если рассматривать активную электрическую проводимость в дендритах (современное представление), то становится очевидным влияние морфологии дендритов: возникает вариация разрядов между всплесками ( англ. bursts) и регулярными формами пульсации , или происходит воздействие на обратное распространение потенциалов действия .
Электрически дендриты могут быть охарактеризованы через их пассивные свойства («скелет»), к которым добавлены (нелинейные) синаптически- и потенциал-зависимые ионные каналы. Пассивный (близкий к потенциалу покоя) скелет дендритов характеризуется удельным мембранным сопротивлением , R m дендритов, которое является относительно высоким (1000—100 000 Ом·см²), откуда следует, что дендритная мембрана является диэлектриком высокого сопротивления. С удельной ёмкостью , C m в приблизительно 1 мкФ /см², мембранная постоянная времени , τ m (которая определяет временной интервал для интеграции синаптических входных импульсов) составляет порядка τ m = R m C m = 10—100 мс . В зависимости от типа и морфологии, площадь поверхности мембранного дендритного дерева колеблется от 20 000 до 750 000 мкм², объём может достигать до 30 000 мкм³, длина — 10 мм (напр., в поясничных мотонейронах ). Внутриклеточная цитоплазма и внеклеточная среда дендритов содержит ионы, способные проводить электрический ток . Дендритная мембрана также способна проводить ток через специфические трансмембранные ионные каналы, но ток, который проходит через мембрану, встречает значительно большее сопротивление, чем вдоль внутренней сердцевины. Кроме мембранных каналов (мембранное сопротивление), дендритная мембрана способна накапливать ионные заряды, то есть функционировать, как конденсатор .
Осевое (продольное) сопротивление дендритной цитоплазмы, R i , колеблется от 70 до 300 Ом·см, что вместе с малыми измерениями отдаленных ветвей предусматривает большое входное сопротивление (импеданс) в дендритах. Увеличение дендритного диаметра с приближением к соме означает большой коэффициент затухания (~100) пикового синаптического потенциала, когда тот распространяется от начала в отдаленной точке дендрита к соме. Мембранное и цитоплазматическое удельное сопротивления обусловливают также внутреннее сопротивление R in в каждой данной точке на дендритном дереве: R in может варьироваться от 1 MОм (толстые и проникающие дендриты) до 1000 MОм (тонкие отростки, как шипики ). Наибольшие значения R in в дендритах показывают, что малое возбуждающее синаптическое изменение проводимости (около 1 нСм) приведёт, локально, к значительному (в несколько десятков мВ) изменению потенциала .
Дендриты, следовательно, являются электрически распределенными «механизмами», поэтому нейроны не являются эквипотенциальными — как их моделировали в классических теориях, — ибо между различными регионами дендритного дерева (и сомы) может существовать десятикратная разница напряжений как результат локального синаптического входа в дендриты.
| Нейрон | Средний диаметр сомы (мкм) | Количество дендритов на соме | Диаметр близких к соме дендритов (мкм) | Количество точек ветвления | Диаметр отдаленных от сомы дендритов (мкм) | Протяженность дендрита א (мкм) | Общая длина дендрита (мкм) |
|---|---|---|---|---|---|---|---|
| Гранулярные клетки мозжечка (кот) | 7 | 4 | 1 | 0 | 0,2—2 | 15 | 60 |
| Звёздчатые амакриновые клетки (резус) | 9 | 1 | 1 | 40 | 0,2—2 | 120 | — |
| Гранулярные клетки зубчатой извилины (крыса) | 14 | 2 | 3 | 14 | 0,5—1 | 300 | 3200 |
| Пирамидальная клетка CA1 (крыса) | 21 | — | — | — | — | — | 11,900 |
| Базальные дендриты | — | 5 | 1 | 30 | 0,5—1 | 130 | 5500 |
| s.radiatum | — | 1 | 3 | 30 | 0,25—1 | 110 | 4100 |
| s.lacunosum-moleculare | — | — | — | 15 | 0,25—1 | 500 | 2300 |
| Клетки Пуркинье мозжечка ( морская свинка ) | 25 | 1 | 3 | 440 | 0,8—2,2 | 200 | 9100 |
| Длинноаксонная клетка бледного шара ( человек ) | 33 | 4 | 4 | 12 | 0,3—0,5 | 1000 | 7600 |
| Клетки Мейнерта зрительной коры (макака) | 35 | — | — | — | — | — | 15 400 |
| Базальные дендриты | — | 5 | 3 | — | — | 250 | 10 200 |
| Апикальные дендриты | — | 1 | 4 | 15 | 2—3 | 1800 | 5 200 |
| Альфа-мотонейрон спинного мозга (кот) | 58 | 11 | 8 | 120 | 0,5—1,5 | 1100 | 52 000 |

Хотя дендриты прилегают ко многим аксонам и др. дендритам, передача сигнала происходит главным образом в синапсах . Синапсы могут находиться непосредственно на поверхности дендрита (т. н. стволовые синапсы), или на дендритных шипиках.

До 1960-х годов считалось, что взаимодействие в нервной системе является односторонним — от аксона к дендритам к соме — и что только аксоны могут быть пресинаптическими. Согласно этому представлению выделяли такие синаптические связи: аксо-дендритные (между аксоном и дендритом), аксо-соматические (между аксоном и клеточным телом), аксо-аксонные (между двумя аксонами) и аксо-шипиковые (между аксоном и шипиком) . Но в 1966 году В. Ролл теоретически предсказал, а его коллеги предоставили физиологические доказательства существования новой и неожиданной синаптической связи, которую они обнаружили в обонятельной луковице — дендро-дендритного синапса. Эти синапсы образуются между дендритами митральной и гранулярной клеток. Оба компонента синапса являются дендритными по своей природе, но ещё более странным было то, что сами эти синапсы являются реципрокными (взаимно обусловленными), поэтому обе клетки действовали как пресинаптические нейроны .
Это открытие было настолько одновременно необычным и важным, что десятилетия после его публикации нейробиологи вспоминали о нём, как о «тихой революции» в понимании нейронов и дендритов: «Новое представление о нейроне [которое вытекает из этого открытия], заключается в том, что дендрит — это отнюдь не пассивная рецептивная поверхность, но также может становиться пресинаптичным, таким, который передаёт информацию в другие нейроны через дендроденритные синапсы» . Другой редкий случай синаптического контакта — это «autapse», когда аксон образует синапс с собственным дендритным деревом, но этот тип связи пока мало понятен .
Синапсы не хаотически распределены на дендритной поверхности. Во многих системах ( пирамидальных клеток гиппокампа или клетках Пуркинье в мозжечке ) источник синаптического импульса выборочно направляется на данный регион дендритного дерева, а не просто случайно распределяется по поверхности дендритного дерева. К примеру, в коре ~79 % всех возбуждающих синапсов находятся на шипиках, а другие — на дендритных стволах, тогда как всего лишь 31 % всех ингибиторных синапсов локализованы на шипиках. Тем более, что шипик с ингибиторным синапсом всегда также имеет возбуждающий синапс: ~15 % всех дендритных шипиков имеют как возбуждающие, так ингибирующие синапсы. Ингибиторные синапсы чаще локализуются на соме или на стволе дендритного дерева . Вследствие такой локализации всего несколько ингибиторных входных импульсов могут шунтировать весь входящий возбуждающий импульс, который собирался дендритным деревом из сотен синапсов апикальных дендритов. Этот феномен называется «ингибиторное шунтирование» .
В целом принято выделять несколько синаптических типов на дендритах :

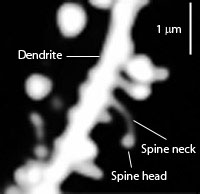
Решающий фактор для способности нейронной сети хранить информацию — количество различных нейронов, которые могут быть соединены синаптически . Одним из главных факторов увеличения разнообразия форм синаптических связей в биологических нейронах является существование дендритных шипиков, открытых в 1888 году Кахалем . Он же впервые выдвинул предположение, что шипики выполняют функцию увеличения поверхности для синаптических контактов с аксонами, находящимися на далёком расстоянии от дендритов: «…дендритные шипики играют главную роль в увеличении поверхности соединений протоплазматического разветвления [то есть, дендритного дерева], выступая, чтобы соединиться с удалёнными нервными волокнами, которые не могут непосредственно соединиться с дендритным отростком» .
В 1904 году Кахаль сделал предположение, что шипики могут хранить электрический заряд , возникающий в результате нейронной активности, тем самым заложив основы понимания синаптической пластичности . Как показали более поздние исследования, шипики расширяют доступность данного дендрита для большего количества аксонов, при этом не слишком увеличивая объём мозга. Хотя большинство шипиков содержат один синапс, существуют шипики с несколькими синапсами. Отношение реального синапса к числу потенциальных синапсов (т. н. коэффициент заполнения ) для пирамидальных нейронов коры составляет 0,26 в неокортексе и 0,22—0,34 в гиппокампе . Это означает, что дендритные шипики играют существенную роль в определении того, какие аксоны образуют контакты с дендритом .
Шипики найдены в разных местах нервной системы , не только в коре головного мозга , а в некоторых случаях, как например, на клетках Пуркинье в мозжечке, они почти похожи на корковые шипики. Впрочем, наиболее важны они в коре головного мозга, поскольку здесь они связаны с 75 % всех синапсов .
Различные регионы дендритных ветвей имеют разную плотность шипиков. К примеру, пирамидальные нейроны CA1 у крысы имеют 30 000 шипиков, 55 % которых сосредоточены в stratum radiatum, а 40 % — в stratum oriens. Плотность шипиков на латеральных ветвях апикального ствола в stratum radiatum достигает, в среднем, 3 на 1 мкм дендрита . Отдаленные дендриты базального конуса имеют подобную плотность шипиков, тогда как в низшем апикальном пучке их плотность значительно ниже. Апикальный ствол в stratum radiatum сам по себе имеет наибольшую плотность шипиков — 7 шипиков на 1 мкм . Наиболее близкие к соме дендриты пирамидального нейрона получают ингибиторные входные импульсы, поэтому эти регионы, так же как и сома, лишены шипиков. У взрослого мозга плотность простых шипиков значительно варьируется от 0,01 мкм³ до более чем 1,5 мкм³ . Тот факт, что различные регионы дендритной ветви имеют разную плотность шипиков, свидетельствует о различиях в соединениях с различными возбуждающими входными синапсами .
Диаметр шипиков колеблется от 0,1 до 0,4 мкм, а длина от 0,4 до 2 мкм. В большинстве регионов мозга шипики имеют два входа — возбуждающий и ингибиторный. Впрочем, в поле CA1 гиппокампа каждый шипик пирамидальной клетки имеет только один синапс — возбуждающий .

Дендритные шипики формируют биохимический и электрический сегмент, где поступающие сигналы вначале интегрируются и обрабатываются. Шея шипика разделяет его голову от остальной части дендрита, тем самым делая шипика отдельным биохимическим и вычислительным регионом нейрона. Подобная сегментация играет ключевую роль в выборочном изменении силы синаптических связей в течение обучения и запоминания .
С шипиками также связаны патологические изменения, поскольку их плотность уменьшается при многих возрастных и психоневрологических заболеваниях, (как деменция , хронический алкоголизм , шизофрения , трисомия ) .
В нейробиологии также принята классификация нейронов на основе существования шипиков на их дендритах. Те нейроны, которые имеют шипики, называются шипиковыми нейронами , а те, которые их лишены — безшипиковыми . Между ними существует не только морфологическое отличие, но и различие в передаче информации: шипиковые дендриты зачастую являются возбуждающими, а безшипиковые — ингибирующими .
Хотя шипики трудно изучать при нормальных условиях, накоплен ряд данных, которые позволяют сделать предположение о существовании различных популяций стабильных и более пластических шипиков in vivo , и эти популяции могут варьироваться с опытом . Существует несколько догадок, что именно эти, динамические шипики , могут участвовать в обучении, тогда как более крупные, стабильные шипики , могут выполнять роль хранителей накопленных воспоминаний . Повторяемая активация малых шипиков ведёт к длительному увеличению в их размерах и восприимчивости к глутамату . И наоборот, незначительная активация AMPA-рецепторов стабилизирует шипики, намекая на двойную роль активации глутамат-рецептора в поддержке структурной пластичности .
В отличие от дендритов, шипики также быстрее реорганизуются и меняются . Их пластичность зависит от актиновых нитей и, скорее всего, коррелирует с обучением . Богатые актином шипики характеризуются особыми морфологическими изменениями, которые связаны с долговременной потенциацией (LTP) и играют ключевую роль в обучении .
| Морфология | Физиология | Синаптология |
|---|---|---|
| Диаметр около сомы : 1—6 мкм | Пассивные свойства дендритов: | Число синапсов на нейрон: 500—200 000 |
| Диаметр на отдалённых концах: 0,3—1 мкм | Мембранное сопротивление ( R m ): 1—100 кОм·см² | Тип І (возбуждающие): 60—90 %; распределены, главным образом на шипиках |
| Средняя длина пути: 0,15—1,5 мм | Осевое сопротивление ( R i ): 70—300 Ом·см | Тип II (ингибирующие): 10—40 %; около сомы, лишь некоторые на шипиках |
| Общая длина дендритов: 1—10 мм | Мембранная ёмкость ( C m ): 1—2 мкФ/см² | |
| Дендритная площадь: 2000—750 000 мкм² | Мембранная временная постоянная ( τ m ): 1—100 мс | Возбуждающий синаптический входной импульс: |
| Дендритные деревья/нейрон: 1—16 | Дендритная пространственная постоянная ( λ): 0,2—1 мм | АМРА : g пик : 0,1—0,3 нс; t пик : 0,3—1 мс |
| Дендритные концы/нейрон: 10—400 | Электротоническая протяженность ( L = x /λ): 0,2—2 | (может увеличиваться с расстоянием от сомы) |
| Дендритные шипики/нейрон: 300—200 000 | Входное сопротивление сомы ( R N ): 1—10³ МОм | NMDA: g пик : 0,05—0,5 нс; t пик : 5—50 мс |
| Плотность шипиков/1 мкм дендрита: 0,5—14 | Входное сопротивление на концах ( R T ) 10²—10³ МОм | |
| Длина шипика: 0,1—2 мкм | Коэффициент затухания напряжения: | Ингибирующий синаптический входной импульс: |
| Диаметр шеи шипика: 0,04—0,5 мкм | сома → конец: 1,1—2 | ГАМК А : g пик : 0,4—1 нс; t пик : 0,2—1,2 мс |
| Диаметр головы шипика: 0,3—1 мкм | конец → сома: 2—15 | ГАМК B : g пик : 0,1—0,3 нс; t пик : 40—150 мс |
| Объём шипика: 0,005—0,3 мкм³ | ||
| Активные свойства дендритов: | ||
| Ca 2+ -каналы (L-, N-, P-типы) — локальный дендритный Ca 2+ -спайк: Ca 2+ -концентрация в шипиках | ||
| Na + -каналы: быстрая активация/инактивация — поддерживает сому → дендритное обратное распространение ПД | ||
| K + -каналы, I A и смешанные токи, I h — увеличение плотности с расстоянием от сомы — «поглотители ударов», нелинейность, временное нормирование |
Пирамидальные нейроны имеют длинный дендритный ствол, который отделяет возбуждающие синапсы на апикальном дендритном дереве от ингибирующих синапсов на базальных дендритах и соме, образуя т. н. открытое поле, в котором дендриты повёрнуты в одном направлении, а сомы в другом. Кроме того, эти клетки характеризуются осевой симметрией и расположены в ряд параллельно друг другу и перпендикулярно к поверхности коры, формируя палисад из клеточных тел и дендритных стволов. Когда оба типа синапсов активны одновременно, ингибирующие синапсы порождают источники тока (из клетки во внеклеточное пространство), а возбудительные — токи стока (из внеклеточного пространства в клетку), заставляя пирамидальную клетку вести себя как микроскопический диполь , окружённый характерным полем, — дендритным потенциалом поля (DFP). Плотно прилегающие друг к другу пирамидальные клетки образуют дипольный слой, наложенные токи которого порождают потенциал локального поля (LFP) нейронных популяций коры, который может быть зарегистрирован с помощью ЭЭГ .
Считается, что пространственная суммация LFP отражает среднее взвешенное возбуждающих и ингибирующих постсинаптических потенциалов, которые являются более медленными, чем потенциалы действия . Хотя уже долгое время считается, что LFP определяется преимущественно синхронизированными дендритными входными сигналами на пирамидальных клетках, но сегодня ещё не ясно, как именно и насколько корреляции в синаптической активности влияют на LFP .
На протяжении нескольких десятилетий — несмотря на их морфологическую сложность, дендритные деревья моделировались при расчётах как единая точка и рассматривались большинством нейробиологов как простые и пассивные механизмы передачи сигналов (изменение мембранного потенциала ) к соме , которая эти сигналы интегрирует в линейный способ и отсылает к аксону , который и генерирует потенциал действия . Тем не менее, вскоре стало очевидным, что пассивные свойства дендритов, наряду с их сложной геометрией, позволяют нейронам интегрировать их входные импульсы в нелинейный способ.
Чтобы понять, какой именно вклад делают дендриты в нейронную обработку информации согласно представлениям дендрита как пассивного механизма, следует иметь в виду, что нейроны генерируют два типа электрических потенциалов: электротонический потенциал и потенциал действия. Электротонический потенциал — это локальный потенциал, который не распространяется активно, а пассивно распространяется вследствие изменения в ионной проводимости (сенсорной или синаптической, которая порождает электрический ток).
Для того, чтобы понять ключевое отличие электротонического потенциала от потенциала действия, необходимое для понимания того, как нейрон передает информацию, — следует различить активный и пассивный путь прохождения потенциалов. Вилфрид Ролл (Wilfrid Rall) в 1960-е годы для описания пассивного электротонического потенциала, который экспоненциально уменьшается проходя вдоль участка мембраны, использовал более широкий термин «расширение» ( англ. spread), обозначавший локальные токи, которые существуют до потенциала действия; этот термин ввиду своей широты также может применяться и к потенциалу действия. Тогда как для описания собственно потенциала действия Ролл использовал термин «распространение» (буквально «размножение») ( англ. propagation), поскольку он точнее соответствует сущности потенциала действия: прохождение последовательными активными процессами воспроизводства самого себя. Итак, электротонические сигналы (напр., ток или потенциал) — это сигналы, которые обусловлены не активными, потенциал-зависимыми свойствами мембраны, а её пассивными свойствами ( RC ). Однако, с открытием первых потенциал-зависимых ионных каналов в дендритах и их способности порождать ПД (см. далее), стало очевидным, что не все дендриты подходят под термин «электротонический», но тем не менее он оказался достаточно полезным для понимания дендритной электрической проводимости и широко используется по настоящее время.
Понимание того, как информация воспринимается и кодируется активными дендритными деревьями, невозможно без понимания модели пассивных дендритов. Более того, при определенных условиях, напр., для малого синаптического входного импульса, потенциалзависимые нелинейности не играют значительной роли, и дендрит действует как пассивная структура.
В 1960-е и 1970-е годы американский математик и нейробиолог Вилфрид Ролл из Национальных институтов здравоохранения в серии статей впервые применил кабельную теорию для описания биофизических и интегративных функций дендритов . В те времена считалось, что дендриты являются настолько электротонически длинными, что любые входящие импульсы к ним не могут существенно влиять на электрические свойства сомы . Тем не менее, соединив математическую кабельную теорию и те незначительные физиологические данные, которые в то время существовали, Ролл убедительно показал, что классические представления о дендритах существенно недооценивают их значительный вклад в нейронные вычисления и передачу информации .
Сведя сложную морфологию дендритов к «эквивалентному цилиндру», кабельная теория Ролла впервые смогла описать электрические и вычислительные свойства пассивных дендритных деревьев с любой сложностью, в частности аналитически вычислить чувствительность напряжения в произвольной точке дендритного дерева к току, поданного в любую другую точку. Это дало возможность оценить амплитуду синаптического потенциала и его зависимость от времени в данном дендритном месте, как и обнаружить, что кабельные свойства дендритов (быстрая зарядка их мембранной ёмкости) фильтруют высокие временные частоты, которые составляют постсинаптические потенциалы (ПСП) , тем самым позволяя дендритам действовать как фильтр низких частот . Теория убедительно доказала, что даже в пассивных дендритах отдалённые синапсы способны существенно влиять на выходной импульс аксона, способствуя значительной деполяризации сомы .
В целом, кабельная теория, разработанная Роллом, продемонстрировала, что комбинация специфической морфологии и электрических свойств мембраны и цитоплазмы (в частности, электротонической структуры) определяет обработку и распространение электрических сигналов в дендритах. Иначе говоря, даже два морфологически идентичных дендритных дерева, но с различными электрическими свойствами, могут иметь совершенно разные вычислительные характеристики.
В пассивном дендритном дереве влияние «возбуждающего постсинаптического потенциала » (ВПСП) на нейронный конечный выходной сигнал зависит от его способности деполяризовать аксон , что, в свою очередь, зависит от а) первичного размера и формы синаптического ответа, и б) от того, насколько кабельные свойства дендритного дерева фильтруют этот ответ в течение его распространения от синапса до аксонного холмика.
Кабельная теория и компартментное моделирование показали, что дендритная морфология играет важную роль в формировании локальных синаптических ответов: отдалённые входные импульсы имеют значительно большие амплитуды локальных реакций, чем подобные синаптические сигналы в более близких к соме регионах . Причиной этого является то, что отдаленные дендриты более сужены (<1 мкм), чем близкие, а потому они создают значительно больший локальный входной импеданс и меньшую локальную мембранную ёмкость, что ведёт к увеличению амплитуды и ускорению прохождения локальных изменений напряжения . С другой стороны, синаптические ответы, возникающие в дендритах, близких к соме, фильтруются существующей здесь большой локальной ёмкостью, что уменьшает их амплитуду и замедляет их временное прохождение. В итоге, быстрые синаптические проводимости в отдаленных дендритных сегментах генерируют бо́льшие и быстрые локальные изменения напряжения, чем такие же проводимости в более близких регионах.
Независимо от локализации на дендритном дереве, в большинстве нейронов единственный синаптический ответ сам по себе не в состоянии деполяризировать мембрану для генерации потенциала действия . В большинстве случаев генерация ПД нуждается в суммации множества ВПСП, взаимодействие между которыми ограничивается теми самыми пассивными свойствами дендритов, что и локальная амплитуда и распространение единичных синаптических ответов. Кабельная теория предполагает , что уровень суммации зависит от относительной изоляции индивидуальных синаптических проводимостей: возбуждающие синапсы, которые находятся в непосредственной близости и совпадают во времени, суммируются сублинейно из-за уменьшения движущей силы (разница между мембранным потенциалом и потенциалом равновесия ионов); тогда как пространственно и временно распределенные синаптические ответы суммируются почти линейно. Эти предположения были подтверждены экспериментально и легли в основу понимания того, как сигналы интегрируются в пассивных дендритах .
Пространственная суммация описывает взаимодействие совпадающих синаптических входных сигналов и зависит от их относительной локализации на дендритном дереве. В пассивном дендрите максимальная (линейная) суммация имеет место только тогда, когда одновременно активированные синапсы настолько пространственно удалённые, что деполяризация, вызванная одним синаптическим импульсом, существенно не уменьшает движущей силы в другом регионе дендрита. Из этого следовало, что одна из возможных функций дендритных деревьев заключается в пространственной изоляции синаптических входных сигналов для дальнейшего усиления их суммации в месте генерации потенциала действия. Эта идея сохраняет свою силу и сегодня, поскольку в ряде исследований было показано, что входные импульсы к нейронам распределены вдоль дендритного дерева .
С другой стороны, тогда как возбуждающие близкие синапсы, когда одновременно активируются, суммируются сублинейно, то синаптические ответы, которые смещены во времени и их изменения проводимости не перекрываются, суммируются линейно. Важным для пространственно-временной суммации в дендритах является то, что постсинаптические потенциалы обычно распадаются с частотой, которая в несколько раз медленнее, чем изменения проводимостей, которые лежат в основе быстрой синаптической передачи сигнала , поэтому уровень и сила синаптической взаимосвязи не являются постоянными и зависят как от кинетики типов рецепторов в синапсе, так и от мембранных свойств и морфологии дендритного дерева.
В целом, как следует из кабельной теории дендритов, следствием взаимодействия между дендритной морфологией и пространственно-временными формами синаптической активации является асимметричный ответ сомы, который возникает в результате последовательной активации синаптических импульсов в дендрите. Как Ролл впервые показал , последовательная активация возбуждающих синапсов на отдалённых регионах дендритов повлечёт большую мембранную деполяризацию, чем подобная последовательная активация, которая бы распространялась от более близких к отдалённым регионам. Это объясняет то, почему нейроны по-разному реагируют на изменения в пространственных формах последовательных синаптических импульсов.
В кабельной теории те же свойства, которые определяют пространственное и временное взаимодействие возбуждающих ответов, также контролируют взаимодействие возбуждающих входных импульсов с синаптическим ингибированием. Нейромедиатором быстрого синаптического ингибирования в мозге млекопитающих является, главным образом, гамма-аминомасляная кислота (ГАМК). ГАМК действует на ГАМК A -рецептор и проводит аниону мембранную проводимость с потенциалом реверсии, близким потенциалу покоя . Поэтому ГАМК может привести либо к гиперполяризующим, или к деполяризующим ответам в нейронах . Способность ГАМК тормозить генерацию потенциала действия лежит в её способности ослаблять совпадающие ВПСП (возбуждающие постсинаптические потенциалы ) через «зажимание» мембранного потенциала ниже порога потенциала действия. Ролл и другие исследователи того времени предвидели, что этот вид шунтирующего ингибирования должен быть более эффективным, чем обычная постсинаптическая гиперполяризация .
Шунтирующее ингибирование придаёт нейрону несколько важных вычислительных преимуществ. Во-первых, поскольку ГАМКергические входные импульсы являются ингибирующими только в течение периода их изменения проводимости, ингибирование является очень точным во времени, тормозя генерации потенциала действия только на несколько миллисекунд, в течение которых ГАМКергическая проводимость активна . Во-вторых, поскольку изменения проводимости значительно более пространственно ограничены, чем обусловленные ими изменения напряжения, то шунтирующее ингибирование обеспечивает эффективное торможение для данного региона дендридтного дерева, при этом сильно влияя на возбудимость в других регионах дендритов . В-третьих, индивидуальные деполяризующие ответы на ГАМК могут (в зависимости от времени и места относительно других возбуждающих входных импульсов) быть как ингибирующими, так и возбуждающими .
Кабельная теория принесла в понимание синапсов то, что синаптические импульсы являются, скорее, изменениями проводимости, чем источниками напряжения , и что их взаимодействие существенно ограничивается дендритной морфологией.
Хотя сегодня уже известно, что дендриты не являются пассивными и способны активно влиять на прохождение синаптического потенциала, кабельная теория пассивных дендритов и поныне используется нейробиологами для изучения биофизических свойств дендритов . А дендриты некоторых типов нейронов, даже те, которые имеют потенциал-зависимые каналы ( интернейроны мозжечка , нейроны-осьминоги слуховой системы , корзинчатые нейроны гиппокампа , гранулярные клетки зубчатой извилины , нейроны грибовидного тела , пирамидальные нейроны обонятельной коры (pyriform cortex) ), характеризуются пассивными свойствами, чем активной проводимостью, а потому лучше могут быть объяснены с помощью кабельной теории .
Хотя уже в 1950-е годы было достаточно много эмпирических данных, которые свидетельствовали в пользу активной роли дендритов в нейронных процессах (напр., открытие распространения потенциала действия по дендритам пирамидальных нейронов ), но они всё равно были в меньшинстве по сравнению с противоположными данными. Именно поэтому открытие потенциал-зависимых ионных каналов в дендритах , , стало одним из наиболее убедительных свидетельств того, что дендриты обладают активными свойствами в обработке входящей информации.
Существует существенная разница между пассивной и активной дендритной проводимостью тока. В случае, когда дендриты пассивны, а сома активна, обработка дендритами возбуждающих постсинаптических потенциалов (ВПСП) минимальна: всё, что произойдет с ВПСП, когда он распространяется к соме, это уменьшение в его амплитуде и увеличение в ширине, как результат межмембранной утечки и фильтрации , . Интеграция потенциалов ВПСП в пассивных дендритах описывается несколькими простыми правилами: потенциалы, которые выходят из мест, электротонически удалённых друг от друга, линейно суммируются (в противном случае ВПСП будут суммироваться сублинейно из-за уменьшения движущей силы синаптического тока и увеличения шунтирования); когда же уже объединённый ВПСП, который достигает сомы, является достаточно большим, возникает потенциал действия (ПД) в аксонном холмике. Далее ПД проходит обратно к соме и вперёд по аксону, где он «передаётся» другим нейронам. При такой модели нейрона длительное раздражение ведёт к повторяющемуся разряду в нейроне с частотой, пропорциональной величине синаптического тока, который поступает к соме .
Однако, данные правила трудно применить к нейрону, который обладает дендритами с активной проводимостью тока, как показали последние исследования . В зависимости от того, какая дендритная проводимость активируется, ВПСП могут ускоряться или затухать до достижения сомы; ВПСП могут суммироваться линейно, сублинейно или суперлинейно. Потенциал действия и другие регенеративные события могут быть инициированы даже в дендритах, образуя разнообразную палитру спаечных форм: от повторяющихся вспышек до пачек (). Такое коренное отличие пассивных дендритов от активных связано с наличием у последних потенциал-зависимых каналов, природа и расположение которых является ключевым для формирования уникального поведения дендритов. Когда эти каналы активируются, токи через них могут изменять амплитуду и временные характеристики синаптического импульса, и даже при определенных условиях генерировать «всё-или-ничего» ( англ. ) регенеративные потенциалы. Следовательно, синаптический импульс, который поступает к соме, будет очень изменённой версией первичного импульса .
Открытие, с использованием микроэлектродов, существования в дендритах крупных кратковременных деполяризаций ( спайков ) приводило к мысли, что дендриты должны содержать ионные каналы . Эти каналы должны были бы открываться в ответ на мембранную деполяризацию и, в свою очередь, оказывать дальнейшую мембранную деполяризацию, которая ответственна за их регенеративную природу. Дальнейшее использование методов локальной фиксации потенциала (пэтч-клэмп) для изучения дендритных свойств в срезах мозга действительно подтвердило существование большого разнообразия потенциалзависимых ионных каналов в дендритной мембране . Вместе с данными, полученными с помощью перфорированного пэтч-клэмпа, эти исследования позволили создать количественные «карты» распределения потенциалзависимых каналов в некоторых типах нейронов. И хотя дендриты имеют те же потенциалзависимые каналы, что и аксон, плотность этих каналов в дендритной мембране — менее чем 10 каналов/мкм², тогда как перехват Ранвье в аксоне содержит до 1000—2000 быстрых натриевых каналов .
Подобно разнообразию в ЦНС дендритных морфологий, так же много разных вариаций ионных каналов в дендритных деревьях. И хотя существует существенная разница между дендритными каналами даже в пределах одного класса нейронов (напр., между корковыми и гиппокамповыми пирамидальными нейронами), накопленные данные позволяют выделить их общие черты и функции. Так, большинство дендритных деревьев (кроме клеток Пуркинье ) содержат некоторые из типов N + -, K + - и Ca 2+ -ионных каналов, как и гиперполяризационно активирующиеся и внутренне-выпрямляющие калиевые каналы . Преимущество потенциал-зависимых ионных каналов в ЦНС состоит в том, что их фундаментальные свойства (разнообразные потенциальные зависимости) и плотность могут быть легко модулированы, что в случае дендритов позволяет нейронам в корне менять способ, которым они хранят и обрабатывают информацию, изменяя саму вычислительную природу нейрона .
Как и в аксонах, в дендритах также были обнаружены Na + -каналы, которые отвечают за генерацию и распространение потенциала действия почти во всех аксонах. В некоторых нейронах ( неокортикальных пирамидальных нейронах , митральных нейронах и пирамидальных нейронах CA 1) плотность Na + -каналов является относительно высокой и постоянной по всему дендритному дереву, что позволяет им влиять на прохождение постсинаптического потенциала , ускорять его , . Наибольшая плотность Na + в дендритах была обнаружена в интернейронах CA1 . Однако в др. типах нейронов (напр., ганглионарных клетках сетчатки , допаминных нейронах чёрной субстанции , гранулярных клетках и т. п.) эта плотность не является такой большой, что объясняет, почему некоторые из этих дендритов не могут сгенерировать потенциал действия .
Относительно однородные свойства активации Na + -каналов порождают быстрый входящий ток, который радикально увеличивает возбуждение дендритов, позволяя им генерировать и распространять потенциал действия так же, как и аксонам, и поддерживать обратное распространение аксонного потенциала действия в дендритные деревья . Дендритные Na + -каналы имеют те же биофизические и фармакологические свойства, как и другие нейронные тетродотоксин -уязвимые каналы . Большинство (>80 %) этих каналов активируются в состоянии мембранного потенциала покоя , а значительная активация Na + -канала начинается, когда деполяризация достигает около 20 мВ. Эти каналы имеют быструю активационную и инактивационную кинетики и основополагающую проводимость отдельного канала в среднем = 15 пСм . Эти каналы обычно модулируются киназами и G-белками , которые способны сильно влиять на потенциал-зависимые свойства дендритов .
Другой тип важных каналов, найденных в дендритах, это K + -каналы, которые являются главными регуляторами возбудимости в нейронах . Потенциалзависимые калиевые каналы (Kv) однородно или менее распределены в отдаленных дендритах по сравнению с сомой. Исключением является их подтип — Kv4-каналы, которые преимущественно локализованы в дендритах . Kv4 субъединицы формируют быстро активированные и инактивированные токи в гетерологичных системах, похожие на K + ток А-типа (IA) в нейронах. Фармакологические , электрофизиологические и иммуногистохимические исследования показали, что K + -каналы А-типа у некоторых типов центральных нейронов более активны в апикальных, радиально наклонённых и базальных дендритах, чем в соме. . Дендриты неокортикальных и CA1-пирамидальных нейронов имеют K + -ионные каналы со свойствами, схожими с K + -каналами с задержанным выпрямлением.
Кратковременные K + -каналы типа А (K A ) были найдены в мозжечковых клетках Пуркинье , где они отвечают за ещё большее усиление сильно угасающих возвратно-протекающих потенциалов действия . Одним из наиболее плотных является присутствие K + -каналов А-типа в пучковых и митральных клетках обонятельной системы, как и в пирамидальных нейронах CA1. Тогда как в таламо-кортикальных нейронах и глубинных клетках неокортекса K A -каналы распределены менее плотно . В целом, эти дендритные калиевые каналы снижают дендритную возбудимость; ограничивают распространение локальных дендритных спайков и Ca 2+ -сигналов, порождённых синаптическими входными импульсами или потенциалами действия обратного распространения; задействованы в реполяризации фазы потенциала действия; ограничивают амплитуду потенциала действия обратного распространения; увеличивают порог для генерации потенциала действия в дендрите, а также контролируют частоту вспышек . Поэтому эти каналы влияют на формы синаптической пластичности, которые зависят от потенциалов действия обратного распространения или локальных спайков.

Дендриты и шипики нескольких центральных нейронов также содержат кальций-активированные калиевые каналы (KCa) . Кальций-активированные калиевые каналы малой проводимости (K Ca 2, или SK) локализованы близко к синаптическим и экстрасинаптическим глутамат -рецепторам, то есть они имеют синаптические функции . В частности, эти каналы уменьшают дендритную интеграцию через ограничение Ca 2+ спайков (плато-потенциалов), вызванных сильным синаптическим входным импульсом . Также, в гиппокампе и миндалине вхождение Ca 2+ через NMDA-рецепторы активирует K Ca 2-каналы, гиперполяризует мембрану и способствует блокированию NMDA-рецепторов магнием (Mg 2+ ), ограничивая дальнейшую активацию. Следовательно, K Ca 2-опосредованная отрицательная обратная связь с NMDA-рецепторами влияет на стимуляцию пластичности Хэбба .
Другой группой K + -каналов, которые, в частности, присутствуют в апикальных дендритах неокортикальных и гиппокампальных CA1-нейронах, являются калиевые каналы внутреннего выпрямления (K ir ) , то есть каналы, которые более легко проводят ток во внутреннем направлении (внутрь клетки), чем в наружном направлении (из клетки). Эти каналы характеризуются внутренним однонаправленным выпрямлением, управляемым внутриклеточной катионной блокировкой . Следовательно, при мембранных потенциалах, более негативных, чем потенциал покоя , K ir -каналы проводят внутренний ток, возвращая мембрану к потенциалу покоя. Однако, при потенциале, более позитивном, чем потенциал покоя, катионы препятствуют гиперполяризации мембраны внешними токами K + . Такие фундаментальные выпрямляющие свойства K ir -каналов являются определяющими для поддержки мембранного потенциала. Среди семи подтипов K ir -каналов K ir 3.x-каналы уникальны тем, что они активируются G-белок-сопряжёнными рецепторами .
Дендриты и шипики нейронов гиппокампа и коры имеют особенно высокую плотность HCN-каналов («гиперполяризационно-активируемые управляемые циклическими нуклеотидами каналы»), в частности каналы HCN1 и HCN2 сосредоточены преимущественно в дендритах. Больше всего HCN-каналов в CA1-нейронах гиппокампа, где их плотность от сомы к ближайшим дендритам возрастает в шесть раз. Но особенно важна их роль в функционировании клеток Пуркинье , где HCN-каналы имеются как в соме , так и в дендритах, поддерживая их тонические импульсы.
HCN-каналы имеют необычные биофизические свойства: они проницаемы как для Na + , так и для K + ; активируются гиперполяризацией (-50 мВ), а деактивируются деполяризацией . Поэтому эти каналы активны в состоянии мембранного потенциала покоя и контролируют его в большинстве нейронов. Катионный ток I h , который проходит через эти каналы, способствует генерации ритмических импульсов (в частности, в сердце и неокортексе ), снижает входное сопротивление клетки, уменьшает возбудимость дендритов и регулирует размер и время протекания синаптических потенциалов к соме . Вследствие уменьшения входного сопротивления (импеданса), I h ослабляет глутаматергические входные импульсы и ускоряет затухание возбуждающих постсинаптических потенциалов, что значительно ограничивает временную суммацию возбуждающих постсинаптических потенциалов и дендритную интеграцию .
Подобно другим активным проводникам, HCN-каналы модулируются внутриклеточными медиаторами. Блокирование I h снижает порог генерации дендритных спайков пачками потенциалов действия обратного распространения, тогда как повышающая регуляция I h снижает возбудимость дендритов пирамидальных нейронов CA1, уменьшая возможность порождения потенциала действия синаптическими входными импульсами .
Кроме того, дендритные I h могут увеличивать точность обнаружения коинциденций (совпадений) отдалённых входных сигналов, а также действовать как фильтр, который оптимизирует апикальные дендриты для низкочастотных осциллирующих входных импульсов . Также ключевую роль I h играет в синаптической пластичности : NMDA-обусловленная LTP увеличивает активность HCN-каналов в CA1-нейронах гиппокампа, тогда как LTD её уменьшает .
Одними из важнейших для активных функций дендритов являются Ca 2+ -каналы ( англ. VGCC, voltage-gated Ca2+ channels), которые, подобно Na + , способны ускорять возбуждающие постсинаптические потенциалы (ВПСП) . Иммуногистохимические и электрофизиологические исследования выявили существование всех подтипов VGCC в дендритных стволах и ряд подтипов в дендритных шипиках многих нейронов ..
Среди Ca 2+ наиболее распространенными являются три типа потенциал-зависимых каналов :
Дендриты некоторых нейронов, такие, как корковые 5 слоя, пирамидальные нейроны CA1 и нейроны в базальных ганглиях , имеют все подтипы каналов HVA (L-, N-, P/Q-, и R-тип) . Относительно медленный, более длительный внутренний ток, который поставляется Ca 2+ -каналами, ещё больше прибавляет возбуждения дендритной мембране и выполняет ряд важных ролей в дендритах: открытие VGCC усиливается синаптическими потенциалами и обратным распространением аксонного потенциала действия, иногда приводя к генерации Ca 2+ -спайков и плато-потенциалов ; локализованные в дендритных шипиках корковых пирамидальных клеток , они являются основой усиления синаптических импульсов ; другие способствуют генерации пачек потенциалов действия, дендритных спайков, а в митральных клетках и нейронах обусловливают кальций-зависимое высвобождение медиатора .
Поскольку внутриклеточный кальций может действовать как вторичный посредник , потенциал-зависимые каналы в дендритах имеют способность соединять электрические сигналы с внутриклеточной биохимией. Поэтому, дендритные Ca 2+ -каналы важны не только для синаптической интеграции, но и синаптической пластичности . Потенциалзависимые Ca 2+ -каналы модулируются G-белками , киназами , фосфатами и липидными модуляторами.
Открытие потенциал-зависимых ионных каналов с различной плотностью и биофизическими свойствами вдоль аксо-сома-дендритной оси впервые показало, что не только сома , но и дендриты играют активную роль в формировании нейронных возбуждений. Поскольку проводимость и распределение потенциалзависимых каналов различается среди различных типов нейронов и дендритных ветвей (напр., апикальные от базальных дендритов), то интеграция входящих синаптических импульсов зависит не только от специфических морфологических и пассивных свойств дендритного дерева, но и от экспрессии и кинетики активной проводимости его ионных каналов .
| Потенциалзависимость | Потенциалзависимость | Кинетика | Кинетика | Кинетика | |||
|---|---|---|---|---|---|---|---|
| Каналы | Активация | Инактивация | Активация | Деактивация | Инактивация | Фармакология | Модулирует |
| Na + | > −45 мВ | > 80 % | < 1 мс | < 1 мс | -1 мс | TTX | PKC |
| Ca T | > −70 мВ | < 25 % | 5 мс | 5 мс | 25 мс | Ni | ? |
| Ca L | > −60 мВ | ~100 % | ~1 мс | < 1 мс | минимальная | PKA | |
| Ca N | > −20 мВ | > 70 % | ~1 мс | < 1 мс | 50 мс | GVIA | G-белок |
| Ca P | > −40 мВ | > 90 % | ~1 мс | < 1 мс | 100 мс | G-белок | |
| Ca R | > −40 мВ | > 50 % | ~1 мс | < 1 мс | 50 мс | Ni | ? |
| K транзиент | > −50 мВ | > 70 % | ~1 мс | < 1 мс | ~7 мс | Киназы | |
| K D | > −50 мВ | > 70 % | ~1 мс | < 1 мс | ~20 мс | αDTX | Киназы |
| K S | > −30 мВ | ~100 % | ~3 мс | < 1 мс | минимальная | Киназы | |
| H | < −50 мв | ~100 % | ~50 мс | 5 мс | никакой | CNT |
Хотя потенциал действия генерируется преимущественно на низкопороговом начальном сегменте аксона (AIS), существует много данных в пользу того, что спайки также могут генерироваться в дендритах (отсюда иногда и различие в названиях: «потенциал действия аксона» и «дендритный спайк»).
Благодаря наличию различных потенциалзависимых каналов с разной кинетикой, дендриты обладают разнообразным репертуаром активных свойств, включая активное обратное распространение аксонного потенциала действия в дендритное дерево . Генерирование в дендритах спайков чаще всего связано с активной проводимостью Ca 2+ - или Na + -каналов, или их обоих . Кроме того, дендритные спайки также порождаются благодаря NMDA -каналам (N-метил-D-аспартат), которые активируются глутамат - нейромедиаторами .
Поскольку локальный порог для генерации этих дендритных спайков значительно выше, чем для потенциала действия в аксоне, то их возникновение обычно требует относительно сильной синхронной синаптической активации, и поэтому может возникать как форма детектора коинциденций (совпадений) . Следовательно, обратное распространение потенциалов действия и дендритные Ca 2+ - и Na + -спайки должны генерироваться различными формами и механизмами синаптической активности.
Рождённые в дендритах спайки не являются однородными в способе, скорости и эффективности их распространения к соме. В некоторых нейронах дендритные спайки успешно распространяются в сторону сомы, почти не теряя в амплитуде . В других нейронах дендритные спайки могут оставаться изолированными в дендритах .
Наличие такого разнообразия дендритных спайков и их вариабельности в различных нейронах позволяет дендритам не только активно влиять на то, как нейроны интегрируют и обрабатывают входные синаптические импульсы, но и на нейронную синаптическую пластичность, обучение и память .
Со времени открытия первых ионных каналов встал вопросо том, зачем на дендритах так много разных ведущих структур, если их влияние не ощутимо на аксонном потенциале действия. Одной из наиболее распространенных идей относительно функций потенциал-зависимых (внутренний ток) каналов в дендритах является то, что они усиливают отдалённые синаптические импульсы. Хотя Вилфрид Ролл доказал, что основываясь на сугубо пассивных свойствах, отдалённые ВПСП не убывают до нуля в соме, как сначала считалось, они тем не менее существенно уменьшаются в амплитуде. Следовательно, Na + - и Ca + -каналы, открытые благодаря ВПСП, противостоят пассивному убыванию импульса через модуляцию или усиление величины ВПСП.
Много каналов в дендритах могут быть активированы подпороговыми ВПСП. Применение методов кальциевой визуализации и пэтч-клемп с прикреплённой клеткой показало, что вызванные стимуляцией сложные ВПСП могут породить в дендритах деполяризацию, достаточную для открытия Na + и низко-пороговых Ca 2+ -каналов. В дендритах существует несколько каналов, на которые могут влиять подпороговые ВПСП: I NaP , I CaT , I h и др. Когда эти каналы активируются, ток через них может изменить форму ВПСП: активация каналов, которые генерируют внутренний ток (I NaP и I CaT ), усилит амплитуду ВПСП, тогда как активация каналов, генерирующих внешние токи (как I А ), ослабит ВПСП. С другой стороны, отключение каналов, которые уже являются активными в состоянии покоя (как I h ), увеличит нейронное входное сопротивление и поэтому увеличит амплитуду ВПСП. В реальности ВПСП меняет состояния активации нескольких каналов одновременно. Общее влияние на форму ВПСП зависит от баланса между каналами, которые генерируют внутренние и внешние токи .
Натриевые и кальциевые каналы , локализованные в шипиках и других отдалённых дендритных отростках, в сочетании с NMDA-рецепторами , обеспечивают уникальный механизм усиления входных локальных синаптических импульсов. В типичном дендритном дереве входящие импедансы на отдалённых ветвях и шипиках растут по сравнению с их низким значением ближе к соме . Следовательно, удалённые возбуждающие синаптические входные импульсы встречают более благоприятные условия для порождения локальных регенеративных потенциалов и значительно больше усиливаются благодаря локальным возбуждающим потенциалзависимым каналам, по сравнению с синапсами на более близких к соме дендритах.
Результаты нескольких экспериментов с пирамидальными клетками CA1 гиппокампа чётко показали, что дендритные I NaP - и I CaT -каналы способны усиливать ВПСП по пути к соме . В этих опытах пэтч-клемп-регистрация целой клетки была сделана на соме, а ВПСП были вызваны через стимуляцию афферентных волокон на удалённых апикальных дендритах. Фокальное приложение антагонистов к ближайшим сегментам апикальных дендритов уменьшило амплитуду ВПСП, в зависимости от блокаторов, на 27 % и 33 %. Тогда как применение антагонистов к соме имело незначительное влияние на амплитуду ВПСП, тем самым подтверждая, что дендритные I NaP и I CaT активно увеличили ВПСП. Более того, моделирование I NaP -каналов показало, что их локальная плотность и пространственное распределение расширяют временной ход усиленных ВПСП, то есть эти каналы могут увеличивать вероятность генерации потенциала действия .
I NaT - и высокопороговые Ca 2+ -каналы также способны усиливать ВПСП. Наиболее разительные примеры усиления сигналов наблюдаются, когда вызванные стимуляцией ВПСП запускают в дендритах Na + -опосредованные потенциалы действия, Ca 2+ -опосредованные регенеративные потенциалы или плато-потенциалы . Хотя, следует отметить, существуют и противоположные экспериментальные данные, которые не так однозначно указывают на важную роль данного усиления для прохождения ВПСП, как и то, происходит ли оно благодаря дендритным или соматическим токам .
Существование дендритных потенциалзависимых каналов и NMDA-спайка являются важными механизмами для междендритных взаимодействий в непосредственной близости синаптических входов. Потенциалзависимые каналы могут менять локальное входное сопротивление и постоянную времени, что в свою очередь существенно повлияет на пространственную и временную суммацию возбуждающих и ингибирующих постсинаптических потенциалов (ВПСП и ИПСП) . Взаимодействие синапсов также может быть чрезвычайно нелинейным: встреча многих ВПСП на одной и те же дендритной ветке в пределах узкого интервала может активировать потенциалзависимые каналы и породить значительно больший ответ, чем если бы они были на разных ветках или произошли вне этого временного интервала . Такой сценарий возможного взаимодействия синаптических потенциалов в дендритах привел нейробиологов к мысли, что дендриты с активными свойствами являются детекторами совпадений ( англ. coincidence detectors) (см. также критические замечания и ответ на них ), то есть они имеют способность «чувствовать» одновременное поступление синаптических входных импульсов в различных точках того самого нейрона. Традиционно выявление коинциденций (совпадений) означало лишь совпадение активации достаточно большого количества входных импульсов для достижения порога генерации потенциала действия. Впрочем, дендриты, как активные механизмы, вводят новые формы выявления совпадений: совпадение спайков во многих дендритных ветвях или совпадение активации многих дендритных регионов.
Одним из таких детекторов совпадений может быть NMDA-рецептор . Из-за его проницаемости для кальция, и благодаря тому, что его блокатор в потенциале покоя Mg 2+ может быть разблокирован постсинаптической деполяризацией, NMDA-рецептор рассматривается как идеальный кандидат для выявления совпадений между пре- и постсинаптической активностью и для перевода её в постсинаптическую кальциевую концентрацию . Следовательно, постсинаптически локализованные NMDA-рецепторы могут обнаружить совпадение в высвобождении глутамата вследствие пресинаптической активности и деполяризацию вследствие постинаптических спайков. Результатом этого является суперлинейный рост концентрации Ca 2+ по сравнению с самой лишь пре- или постсинаптичною активностью. Свидетельства такого выявления совпадений на основе NMDA-рецептора были обнаружены в гиппокампе и неокортексе . Подобное дендритное выявление совпадений было обнаружено и для потенциалзависимых каналов. В отдаленных синапсах на апикальных дендритах пирамидальных нейронов 5 слоя, объединение постсинаптического потенциала действия и синаптического входного импульса может вызвать весьма нелинейное усиление дендритных потенциалов действия обратного распространения благодаря привлечению потенциалзависимых Na + -каналов .
Хотя гипотеза о дендритах как детекторах совпадений является темой жарких дебатов между учёными, существование такого механизма имеет ряд экспериментальных подтверждений. К примеру, в пирамидальных нейронах CA1 синапсы апикального пучка не проявляют синаптического масштабирования и поэтому, вероятно, влияют на потенциал действия или суммирование с синаптическими входными импульсами от коллатералей Шафера (связка волокон, которая идёт от поля CA3 до поля CA1 гиппокампа), или через генерацию дендритных спайков . Как показали эксперименты и симуляции, отдалённые дендритные спайки надежно не распространяются от апикального пучка до главного апикального дендрита . Эта невозможность распространения дендритного спайка связана со значительным падением входного импеданса в точках, где малые ветви соединены с большими дендритами. Когда же колатераль Шаффера и перфорантный путь ( англ. perforant path) активируются вместе, дендритные спайки могут распространяться надежно до сомы. И наоборот, ингибирование, нацеленное на апикальный дендрит, делает невозможным распространение дендритного спайка . Итак, синаптический входной импульс к апикальному дендриту может открыть или закрыть «ворота», регулирующие влияние удалённо генерируемых дендритных спайков на начало аксонного потенциала действия.
Один из наиболее ярких экспериментальных примеров, который свидетельствует в пользу существования дендритного детектора совпадений, был обнаружен в пирамидальных нейронах 5 слоя: возбуждающие синаптические входные импульсы на удалённом апикальном пучке, совпадая с потенциалом действия обратного распространения, генерируют большой (10 мск и больше) Ca + спайк ( англ. BAC firing), который, в свою очередь, распространяется в сторону сомы и побуждает аксон породить пачковые ( англ. burst) натриевые потенциалы действия. Натриевый потенциал действия обратного распространения служит как «связывающий» механизм для специфической комбинации входных импульсов в дендритном дереве . Этот механизм позволяет выявить совпадение активаций синаптических входных импульсов к двум главным сегментам дендритного дерева и, следовательно, может быть вовлечён в одновременную активность ( синхронизацию ) в различных слоях коры.


Кабельная структура дендритного дерева, как следует из кабельной теории и экспериментальных данных, приводит к затуханию синаптического потенциала. В частности, длинные тонкие дендриты, которые имеют большое осевое сопротивление, существенно ослабляют потенциал в подпороговой фазе . К примеру, затухание пиковой амплитуды ВПСП в ходе их распространения от места возникновения до сомы может быть более чем стократным для наиболее удаленных синапсов в корковых пирамидальных нейронах L5 . Частично это связано с низкочастотной фильтрацией, что проявляется в быстрых скачках напряжения в дендритах и что также может привести к существенным замедлениям электрической проводимости . Из-за такого затухания дендритного напряжения синапсы в разных точках дендритов не являются одинаково эффективными в их воздействии на аксонный потенциал действия. Этот факт породил понятие «дендритной демократии» , когда все синаптические входные импульсы любого нейронного типа могут быть одинаково «услышаны» и почти мгновенно вместе обработаны сомой.
Затухание заряда в длинных и тонких дендритах пирамидных клеток существенно уменьшает соматическую амплитуду ВПСП, которые возникают в синапсах у отдалённых дендритов, по сравнению с ВПСП, возникающими в ближайших синапсах с той же самой синаптической проводимостью, тем самым делая эти нейроны скорее «дендритно не-демократическими» . Но совсем другая ситуация с клетками Пуркинье , чьи шипиковые веточки относительно короткие и непосредственно соединены с более толстыми главными дендритами. Следовательно, равная синаптическая проводимость на отдалённых и близких шипикових ветвях клетки Пуркинье предопределяет очень похожие соматические амплитуды ВПСП . Это очередной пример влияния морфологии дендритов на их проводимость и целостное нейронное поведение: дендритная геометрия клеток Пуркинье по своей природе является более «демократичной», чем, напр., пирамидальных клеток. «Дефицит демократии» пирамидальных клеток частично компенсируется связанным с расстоянием масштабированием синаптической проводимости в апикальных дендритах нейронов CA1 .
Однако этот тип компенсирующего синаптического масштабирования не был найден в базальных и апикальных дендритах нейронов 5 слоя коры . Одно из решений этой ключевой для понимания активной природы дендритов проблемы заключается в том, что «не вполне демократические» дендриты неокортикальных пирамидальных нейронов — в силу сложности информации, которую они обработают — состоят из относительно независимых вычислительных субъединиц, которые сигмоидально модулируют их синаптические входные импульсы до глобального суммирования и каждая из которых способна порождать дендритный спайк. Приведут ли эти спайки к аксонному потенциалу действия, будет зависеть от интеграции единичных ответов этих субъединиц .
Один из возможных компромиссов между «демократией» и «независимостью» в дендритной обработке информации недавно был найден при изучении не пирамидальных клеток, а т. н. координатных клеток (звёздчатых нейронов энториальной коры ) и заключается в том, что если дендриты имеют сильное влияние на соматический мембранный потенциал («демократия»), тогда, из-за двусторонности электротонического соединения (существование градиента напряжения между дендритами и сомой, как результат потенциала действия обратного распространения), сома будет сильнее влиять на дендритные процессы (потеря «независимости» — способности самостоятельно от других дендритов интегрировать свои входные сигналы), вырождая их локально суммированную информацию. Иначе говоря, увеличение дендритной независимости ведёт к уменьшению их влияния на сому и, как следствие, к уменьшению синаптических входных сигналов на сами дендриты.
Дендритные спайки могут быть порождены кластерными или распределенными входными импульсами к одной дендритной ветви. Однако входные импульсы, распределённые на многих дендритных ветвях, будут менее эффективны, поэтому они, скорее всего, должны группироваться в кластеры . По такому сценарию соседние входные импульсы на той же самой ветке суммируются сигмоидально, тогда как слишком отдалённые входные импульсы к различным ветвям суммируются линейно. Подобная пространственная сегментация обработки импульсов в дендритах не поддерживает идею о глобальной суммации и вместо этого позволяет говорить о пирамидальном нейроне как двухслойной «нервной сети», где дендриты предстают как «нейроны в нейронах»: на первом слое синаптические входные импульсы интегрируются индивидуальными ветвями, сигмоидальными субъединицами (что соответствует длинным, тонким дендритам клетки), а на втором слое выходные импульсы этих субъединиц суммируются в главном дендритном пучке и соме, прежде чем достичь необходимого порога генерации потенциала действия .
Эта идея также может быть расширена: пирамидальный нейрон рассматривается как многослойная сеть, где интеграция отдельно происходит в апикальном пучке, апикальном наклонении, а базальные дендритные ветви действуют как промежуточные слои . Эти идеи недавно нашли ряд экспериментальных подтверждений (См. пример с пирамидальными нейронами CA1 в предыдущем разделе). Так же было открыто, что распространение дендритных спайков поддерживается синаптически активностью на заднем фоне в пирамидальных нейронах 5 слоя .
Тем не менее, двухслойная модель нейрона описывает только пространственные группировки синаптических входных импульсов, оставляя вне поля зрения, как нейрон обрабатывает темпоральные формы входных импульсов. Сейчас предложена только одна компартментная модель, которая способна объяснить темпоральную интеграцию импульсов дендритами. Согласно этой модели, ответ дендритных сегментов может быть описан как нелинейная сигмоидальная функция одновременно и степени временной синхронности, и пространственного группирования синаптических входов. Тогда нейрон предстаёт как многослойная сеть: дендриты выборочно усиливают ответы к релевантным пространственно-временным входным спайкам, тем самым действуя в различных интегративных режимах как многослойный детектор совпадений . Эта модель также согласуется с экспериментальными данными, которые показывают, что в различных поведенческих состояниях могут быть задействованы различные формы нейронной интеграции .
Кластеризация и недемократичность упомянутых нейронов обусловлена сложной морфологией их дендритов, существованием активных токов и локальных спайков, которые по-разному влияют на интеграцию постсинаптических потенциалов (ПСП). Значительно более демократическими являются нейроны, дендриты которых имеют пассивные кабельные свойства, как к примеру, пирамидальные и не-пирамидальные нейроны CA3, вставочные нейроны мозжечка, гранулярные клетки зубчатой извилины . Моделирование и исследование in vivo пассивных дендритов свидетельствуют, что синаптический импульс к проксимальному дендриту вызовет соматический ПСП, который только на 10—12 % больше, чем ПСП, вызванный синапсом на наиболее удаленном дендрите .
То есть, для пассивных дендритов, в отличие от активных, положение синапса не имеет значительного влияния на соматический ПСП, и все синапсы имеют равный «голос» в соме. Механизмом такой демократизации является т. н. «пассивная синаптическая нормализация» входных импульсов, которая трансформирует локальные высокоамплитудные ПСП — которые широко рассеиваются по клетке, но имеют ограниченный диапазон — в низкоамплитудные колебания мембранного потенциала близко к соме .
Известный постулат Дональда Хэбба , который является основой классической теории синаптической пластичности, трактует все синапсы равными и игнорирует любой вклад дендритов в синаптическую пластичность. Как он писал в своём учебнике по психологии: «…[Ф]ункцией дендритов является получение возбуждений от других клеток… Дендритная проводимость является медленной и неэффективной, считается, что они являются примитивными структурами, а непосредственное возбуждение тела клетки — это эволюционное совершенствование, которое позволяет более эффективную проводимость» . Подобно Хэббу, другие нейробиологи долгое время считали, что дендриты выполняют лишь соединительную функцию и только передают информацию от синапсов к соме , не играя какой-либо значительной роли в синаптической пластичности, изменении входных импульсов и вычислениях нейрона.
Коренное изменение в представлении о природе и функции дендритов была связана с теоретическими трудами Вилфрида Ролла и его учеников и коллег, которые показали, что даже пассивные дендриты имеют значительное влияние на электрические свойства сомы. Впрочем, с вычислительной точки зрения, пассивные дендриты могут выполнять лишь несколько элементарных операций: фильтр низких частот , арифметика с насыщением и похожие на умножения взаимодействия между синаптическими входными импульсами . Однако значительно более богатый и сложный репертуар нелинейных и нестационарных операций появляется, если дендритные деревья имеют потенциал-зависимые мембранные проводимости.
Кроме уже указанных интегративных свойств, активные дендриты обладают сложным и пока что мало понятным аппаратом для синаптической пластичности . За последние несколько десятилетий был найден целый массив свидетельств дендритной пластичности, в том числе синаптическая (гомо- и гетеросинаптическая) и внутренняя пластичность и гомеостатические механизмы, которые часто действуют локально и параллельно друг с другом, регулируя активные свойства дендритов, тем самым влияя не только на дендритные вычисления, но и на формирование памяти и обучения на субклеточном уровне .
Кроме того, благодаря сложной морфологии и существованию многочисленных потенциал-зависимых ионных каналов , дендриты превращают одиночные нейроны в мощные функциональные вычислительные микросхемы, которые способны выполнять операции, которые раньше считались возможными только для популяции нервных сетей. В частности, в течение последних двух десятилетий было открыто, что дендриты способны выполнять синхронизацию и классификацию входных синаптических сигналов , выявлять направление движения (направленная избирательность) в зрительной системе , параллельно вычислять различные потоки информации локализовать источник звука в слуховые системе , а дендритные шипики даже могут быть настроены на разные частоты и интенсивности тонов .
Все эти вычислительные свойства дендритов делают даже индивидуальный нейрон способным к активной обработке сложной информации и её хранению, тем самым решая ряд классических проблем в нейробиологии , которые долгое время не могли быть решены, поскольку большинство подходов к ним основывалось главным образом на точечной модели нейрона, без учета активной роли дендритов .
Одним из наиболее убедительных свидетельств относительно активной и важной роли дендритов в нейронных процессах являются нейродегенеративные, возрастные и психические расстройства, которые с ними связаны. С увеличением данных об интегративных и пластических функциях дендритов, также растёт понимание того, почему даже небольшие структурные изменения в дендритах способны привести (или сопровождать) к значительным нарушениям в нормальном функционировании мозга .
В 1974 году Доминик Пурпура (Dominick Purpura) в своей уже классической статье в « Сайенс » выдвинул гипотезу, что открытые в те времена аномалии дендритных шипиков (дисгенез) лежат в основе некоторых из типов умственной отсталости . Эта статья, вместе с другими положила начало детальному изучению дендритов и шипиков и их связи с психоневрологическими заболеваниями. С тех пор была открыта сильная корреляция между дендритной патологией и умственной отсталостью, в частности такими заболеваниями, как аутизм , синдромы Дауна , Ретта , Мартина — Белл , Вильямса и Рубинштейна — Тейби .
В целом, эти и другие возрастные расстройства характеризуются уменьшением дендритной длины, сокращением форм разветвлений и количества шипиков. Те же шипики, которые остаются, часто является очень длинными и тонкими . Понятно, что выполняя важные биологические и вычислительные функции, так кардинально изменённые дендриты не могут функционировать нормально. Вместе с тем, изменённая морфология может и не быть первичной причиной расстройств, зато выступать как компенсационное или вторичное изменение, связанное с другой, более первичной патологией. К примеру, многие из указанных изменений дендритной морфологии могут возникать вследствие (потери возможности проведения сенсорного возбуждение от периферии к центру) .
Первые свидетельства о изменениях в морфологии дендритов, обусловленных патологическими нарушениями, были получены при исследовании поражений, вызванных деафферентацией и, как следствие, потерей синаптических входных импульсов. Было обнаружено, что эти поражения привели к полному уменьшению, деформации и дезориентации дендритов в мозжечковых клетках Пуркинье . Обусловленные поражениями дендритные изменения (существенное сокращение длины отдаленных дендритов) также были обнаружены в гранулярных клетках в зубчатой извилине как следствие деафферентации энториальной коры. С другой стороны, был обнаружен и обратный эффект — распускание базальных дендритов гранулярных клеток гиппокампа, вызванное длительной эпилептиформной активностью .
Довольно давно известно, что потеря нейронов и синапсов в определенных регионах мозга является одним из последствий нормального старения у здоровых взрослых. Кроме того, в процессе старения были обнаружены также изменения в дендритной структуре и количестве шипиков , хотя степень и специфика этих изменений пока не известны. Поэтому сегодня ещё трудно связывать эти морфологические изменения с когнитивными нарушениями.
Кроме того, ряд исследований показал, что значительные изменения в морфологии дендритов, шипиков и синапсов характерны для болезни Альцгеймера и шизофрении . Бета-амилоидний пептид , который может вести к образованию амилоидных бляшек и который связан с болезнью Альцегймера, может блокировать А-тип потенциал-зависимого калиевого канала в дендритах пирамидальных клеток. Особенно уязвимы для разрушительного воздействия бета-амилоидов калиевые каналы тонких ветвей наклонённых дендритов, в результате чего может возникать ухудшение когнитивных функций .
Накопление особых протеинов (т. н. тельца Леви ) в дендритах часто характерно для болезни Паркинсона . Однако сейчас трудно говорить о функциональных последствия таких изменений и степени их связи с данными заболеваниями.
| Годы | Методы и технологии | Результаты |
|---|---|---|
| 1870-е | Метод Гольджи | Дендритная морфология |
| 1930-е | Внеклеточного поля | медленные волны коры |
| 1950-е | Внутриклеточная регистрация | возбуждающий постсинаптический потенциал (ВПСП); за отдельными исключениями, пассивные дендриты |
| 1959 | Кабельная теория | Дендриты влияют на электрические свойства сомы |
| 1960-е | Количественная микроанатомия | Дендритные параметры и измерения |
| 1970-е | Междендритная регистрация | Дендритные потенциалы действия |
| 1980-е | Компартментное моделирование | Активные и пассивные свойства |
| 1980-е | Молекулярная биология | Молекулярное разнообразие каналов |
| 1990-е | Флуоресцентная микроскопия | [Ca 2+ ] i вхождения в дендритах, шипиках |
| 1990-е | Пэтч-кламп дендритов . | Активность и интегративность каналов |
| 1997 | Двухфотонная микроскопия | Регистрация единичного дендрита, кальциевые импульсы и детектор совпадений |
| 1999 | Потенциалзависимые красители | Одновременная регистрация сомы и дендрита, аксонный ПД, дендритний спайк и bAP |
| 2006 | Высвобождение глутамата (glutamate uncaging) | Дендритная пластичность |
| 2002—2007 | Динамический кламп (dynamic clamp) | Двухуровневая структура, «дендритная демократия» и дендритные вычисления |
| 2010—2013 | Моносинаптичне отслеживания с использованием вирусных векторов; GCaMP | Кластеризация шипиков; Визуализация дендритного потенциала действия, LTP и направленная избирательность |
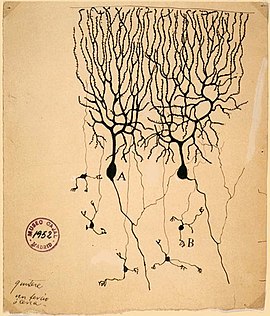
Первое подробное описание дендритов (или «протоплазматических отростков», как они сначала назывались) было сделан Камилло Гольджи в 1873 году. Однако, он не понял их функции, считая, что они выполняют только питательную роль для нейрона . Первым, кто начал трактовать дендриты как самостоятельные функциональные единицы, был Сантьяго Рамон-и-Кахаль , который, использовав разработанный Гольджи метод окрашивания нервных тканей, предложил нейронную доктрину, согласно которой дендриты представали как место синаптических контактов между нейронами и выполняли функцию приёма и передачи синаптических импульсов. Когда функции и типы дендритных деревьев были изучены более детально, Вильгельм Хис ( англ. Wilhelm His) в 1889 году заменил термин «протоплазматические отростки» на «дендриты» .
Позже Рамон-и-Кахаль для объяснения механизмов нейронной обработки информации предложил концепцию «динамической поляризации», согласно которой информация протекает в одном направлении: синапс → дендрит → сома → аксон . Гипотеза, которая лежала в основе такого предположения, заключалась в том, что, для выполнения интегративных функций нейроном, в нём должна происходить градуировочная суммация ингибирующих и возбуждающих импульсов, иначе нейроны и дендриты будут функционировать только как реле , не изменяя и не добавляя информацию в процессе её передачи.
До 1930-х годов изучение дендритов было главным образом анатомическим, и лишь когда методы исследования аксонов (полученные на основе изучения периферических нервов ) были применены к изучению электрических сигналов в коре головного мозга , дендриты начали изучаться электрофизиологически . Стимулирование зрительного нерва или поверхности коры вело к появлению отрицательного потенциала (несколько милливольт) на поверхности, что было слишком медленным, чтобы быть следствием аксонных потенциалов действия , которые регистрировались в изолированных периферических нервах . Тогда Джордж Бишоп (George H. Bishop) с коллегами , используя большие электроды для регистрации в дендритных слоях зрительной коры, сделали предположение, что эти отрицательные волны поверхности коры являются непроводящими, постоянными потенциалами, которые впоследствии Экклс трактовал, как синаптические потенциалы в дендритах. Основываясь на этих догадках и собственных исследованиях, Бишоп пришел к выводу, что «главная и наиболее характерная функция нервных и других возбуждающих тканей осуществляется с помощью градуировочных реакций» . Поскольку такое представление электротонического распространения реакций в дендритах согласовывалось с идеями динамической поляризации Кахаля и нейронной интеграции Чарльза Шеррингтона , оно было наиболее распространено (за отдельными исключениями ) среди нейробиологов вплоть до начала 1960-х годов .
С появлением внутриклеточной регистрации с помощью микроэлектродов, во многих типах нейронов были открыты возбуждающие и ингибирующие постсинаптические потенциалы (ВПСП и ИПСП). Также были определены свойства синапсов (потенциал равновесия) и измерены ключевые параметры постсинаптической клетки (мембранные временная и пространственная постоянные). Накопленная новая информация позволила поставить ряд вопросов относительно того, как осуществляется интеграция пассивных синаптических потенциалов в дендритах, их пространственная и временная суммация. Было выдвинуто несколько предположений , что дендриты имеют более примитивную мембрану, чем аксоны, и поэтому не имеют возможности для активной электрической проводимости. Соответственно, считалось (на основе исследований мотонейронов и сенсорных рецепторов потенциал действия возникает только в аксонном холмике, как следствие алгебраической суммации ВПСП и ИПСП, поступающих в разные части нейрона.
Поскольку основанные на тогдашних электрофизиологических данных расчёты для мотонейронов показали, что пространственная постоянная (λ) является относительно малой, то ряд исследователей, особенно Джон Экклс , посчитали, что ВПСП из синапсов в отдалённых регионах дендритов не имеют значительного влияния на электрическое поведение нейронов и генерацию потенциала действия, учитывая быстрое снижение их амплитуды ; только синапсы на более близких к соме дендритах могли участвовать в нейронной активности.
Поэтому для многих нейробиологов стала большой неожиданностью опубликованная в 1957 году статья малоизвестного Вилфрида Ролла (род. 1922), бывшего постдока Экклса, после которой появилась серия других его статей, которые изменили не только представление о дендритах, но и о нейронах . После отличного обучения на физическом факультете Йельского университета, с началом войны он был привлечен к аналитической работе в Манхэттенском проекте . После войны, работая как постдок в Чикагском университете , Ролл получил навыки в электрофизиологии и участвовал в экспериментах с аксоном кальмара , которые проводил известный американский биофизик Кеннет Коул ( англ. Kenneth Stewart Cole), и которые в результате привели к открытию потенциала действия . С начала 1950-х годов он продолжил свои исследования в лаборатории Экклса в Новой Зеландии , где с его группой изучал передачу сигналов в мотонейронах и использование микроэлектродов для регистрации этих сигналов. Поработав некоторое время в лаборатории Бернарда Каца в Лондоне, он вернулся в США, где начал работу на математическом отделении Национальных институтов здоровья .
В 1957 году Экклс и другие исследователи опубликовали результаты исследований мотонейронов, которые показывали распространение потенциалов, вызванных инжекцией тока в тело клетки. Фазы распада потенциалов, считал Экклс, выражаются лишь одной экспонентой , так, как в простой схеме сопротивления и ёмкости, которыми моделируется мембрана сомы. Проанализировав данные Экклса и его группы, Ролл обнаружил, что регистрируемые ими кратковременные потенциалы были значительно медленнее, чем ожидалось, и скорее всего это связано с тем, что ток идет в дендриты, а следовательно временная и пространственная постоянные нейронов значительно больше. В том же 1957 году он опубликовал короткую заметку в « Сайенс », где показал, что зафиксированные Экклсом кратковременные потенциалы больше согласуются с моделью сомы, к которой присоединен длинный цилиндр, которым является дендритное дерево .
Когда Ролл осознал, что дендриты играют значительную роль в электрической проводимости нейронов, он начал детальное изучение геометрии разветвлений дендритов мотонейронов, что позволило ему применить к ним кабельную теорию , сведя всё многообразие дендритных деревьев к одному цилиндру. Это не только упрощало изучение дендритов, но и позволяло открыть их новые свойства: в частности, Ролл предсказал, что синапсы на удаленных дендритах также влияют на соматическую деполяризацию. Однако все статьи, которые он посылал в журналы, отклонялись редакторами, с аргументацией, что найденное Роллом кратковременное снижение потенциала было связано с особенностями технических приборов и не имело существенного значения. Впрочем, на нескольких статьях Ролл увидел комментарии и изменения, характер которых вполне свидетельствовал, что их читал Экклс. Тогда Ролл опубликовал свои теоретические статьи в новом, малоизвестном журнале «Экспериментал Нэуролоджи», в котором Экклс не был рецензентом .
Только через десятилетия, когда накопились новые данные, теории Ролла получили подтверждение и признание, а его кабельные уравнения и математические модели дендритов заложили основу новой области нейронаук — вычислительной нейронауки ( англ. computational neuroscience). Его последующие научные работы были очередным ярким примером того, как в биологии могут эффективно сочетаться математика и эксперимент, а теория может иметь не менее важное значение, чем практика. Многие из проблем в изучении дендритов, которые затем сформировались в отдельные темы, были так или иначе подняты, предвосхищены или решены Роллом. Поэтому, «история кабельной теории и дендритов является во многом историей одного человека — Вилфрида Ролла» .
Впрочем, для того, чтобы вычислительные модели могли объяснить дендритные свойства, не хватало данных по их детальной анатомии и физиологии. Именно поэтому в 1970-е годы новую страницу в изучении дендритов открыла количественная микроанатомия, то есть детальное и точное измерение дендритных диаметров и длины ветвей. Благодаря кропотливой работе анатомов, физиологов и математиков были получены первые детальные параметры корковых нейронов , мотонейронов , клеток Пуркинье и т. п. Благодаря появлению вычислительной техники и более-менее гибких языков программирования, полученные микроанатомические данные легли в основу вычислительных моделей, на основе которых был открыт целый ряд новых дендритных свойств .
Хотя в 1950-е и ранние 1960-е годы господствующим было представление, что дендриты являются пассивными отростками нейронов, которые просто интегрируют возбуждающие и ингибирующие импульсы, накапливались единичные свидетельства, что в дендритах также может распространяться потенциал действия . Впервые зафиксировал его в 1951 году выдающийся китайский нейрофизиолог , который тогда работал в Йельском университете и имел ещё более драматическую судьбу, чем Ролл . В опубликованной в том году статье он сообщил о том, что дендриты могут возбуждаться под воздействием электрической стимуляции и способны генерировать потенциалы действия, которые отличаются от аксонных тем, что не являются «все-или-ничего» потенциалами . В следующих восьми своих публикациях он подтвердил свои находки и даже выдвинул радикальную догадку, что синапсы на дендритах, в отличие от сомы, связаны с сознанием , восприятием и мышлением .
Другой важным догадкой Чанга было признание за дендритными шипиками (он их называл «почечками») функции ограничения синаптической возбудимости, поскольку они предстают как механический барьер, предотвращающий достижение синаптическими выступами ствола дендрита . Шипики, благодаря их высокому омическому сопротивлению, связанному с их слишком тонкими «стеблями», должны замедлять и ослаблять синаптические возбуждающие импульсы, тем самым играя активную, а не пассивную роль в синаптической интеграции . Именно на основе этих результатов Чанга в 1970-е годы возник новый интерес к шипикам и их роли в обучении и запоминании.
Также в 1950-е годы годах ряд исследователей сообщал о регистрации антидромических потенциалов действия и дендритных спайков , что должно было бы склонить больше нейробиологов к признанию активной роли дендритов. Тем не менее, лишь с конца 1980-х — начала 1990-х годов учёные начали постепенно склоняться к мысли, что дендриты не только передают информацию, но и меняют её и хранят. Наиболее однозначно существование дендритных спайков было показано в серии статей Грега Стюарта и Берта Сакмана в течение 1993—1998 годов , которые использовали цело-клеточные пэтч-электроды для регистрации одновременно соматического потенциала действия и дендритного спайка. Это были первые прямые свидетельства существования потенциалзависимых ионных каналов в дендритах, которые служат для генерации и поддержания потенциалов действия.
1990— 2000-е годы могут быть названы периодом расцвета исследований дендритов. Стремительный прогресс технологий, молекулярной биологии и вычислительных техник привёл к быстрому появлению новых открытий, связанных с дендритными вычислениями и пластичностью, как структурной, так и функциональной.