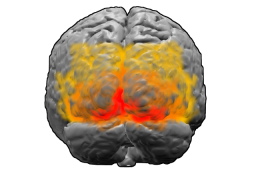
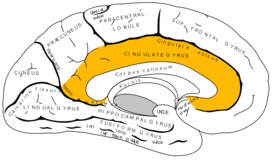
Переходная новая кора
- 1 year ago
- 0
- 0

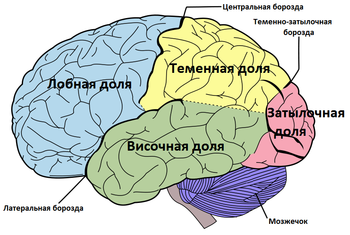
Премоторная кора ( англ. Premotor cortex , PMC ) — область моторной коры , расположенная в задних частях верхней и средней лобной извилины и в передних частях прецентральной извилины , не занятых первичной моторной корой . Соответствует латеральной части цитоархитектонического поля Бродмана 6 . Вместе с дополнительной моторной областью ( англ. Supplementary motor area) составляет так называемую вторичную моторную кору ( англ. Secondary motor cortex).
Функции премоторной коры разнообразны и включают в себя контроль движений туловища, планирование движений, контроль движений на основе сенсорной информации и другие. Различные области премоторной коры обладают отличными свойствами и, вероятно, отвечают за разные функции.
Кортикоспинальные волокна, отходящие от премоторной коры, составляют примерно 30 % волокон пирамидного тракта .
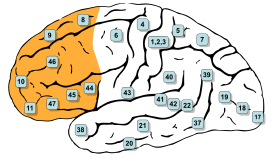
Премоторная кора занимает латеральную часть цитоархитектонического поля Бродмана 6 . В медиальной части поля Бродмана 6 располагается дополнительная моторная область .
Премоторная кора отличается от первичной моторной коры ( поле Бродмана 4 ) двумя анатомическими особенностями. Во-первых, первичная моторная кора содержит гигантские пирамидальные нейроны под названием клетки Беца в V слое, тогда как в премоторной коре пирамидальные нейроны меньше по размеру и их меньше. Во-вторых, первичная моторная кора является агранулярной: в ней нет IV слоя гранулярных клеток, тогда как в премоторной коре этот слой присутствует, хотя он и небольшой.
Также премоторная кора отличается от префронтальной коры , который находится перед ней, наличием полностью сформированного внутреннего зернистого IV слоя в поле 46. Таким образом, премотороная кора анатомически отличается от агранулярной первичной моторной коры и гранулярной, шестислойной префронтальной коры.
Премоторная кора разделяется на несколько субрегионов на основании цитоархитектоники (выделении клеточных слоёв при изучении под микроскопом), гистохимии (изучении химического состава тканей), анатомических связей с другими областями мозга и физиологических свойств .
Связи премоторной коры с другими областями мозга разнообразны и включают как афферентные (входящие), так и эфферентные (исходящие) нервные пути с первичной моторной корой, дополнительной моторной областью, теменной долей , префронтальной корой, а также с подкорковыми структурами, включая спинной мозг , полосатое тело , таламус и другие области.
В настоящее время премоторная кора разделяется на четыре секции (зоны) . В первую очередь выделяется верхняя (дорсальная) и нижняя (вентральная) премоторная кора. Каждая из этих областей дополнительно разделяется на участки, лежащие ближе к передней части мозга (ростральная премоторная кора) и лежащие ближе к задней части мозга (каудальная премоторная кора). Для этих областей в англоязычной литературе применяются следующие аббревиатуры: PMDr — премоторная дорсальная ростральная кора ( англ. Premotor dorsal, rostral), PMDc — премоторная дорсальная каудальная кора ( англ. Premotor dorsal, caudal), PMVr — премоторная вентральная ростральная кора ( англ. Premotor ventral, rostral), PMVc — премоторная вентральная каудальная кора ( англ. Premotor ventral, caudal). Ряд исследователей использует другую терминологию. Поле 7 или F7 ( англ. Field 7 , англ. Field F7) соответствует премоторной вентральной ростральной коре (PMDr), поле F2 — премоторной дорсальной каудальной коре (PMDc), поле F5 — премоторной вентральной ростральной коре (PMVr), поле F4 — премоторной дорсальной каудальной коре (PMVc).
Следует отметить, что вышеуказанные области премоторной коры были впервые выделены в мозге обезьян и изучены в основном на обезьяньем мозге.
Премоторная дорсальная каудальная область (PMDc, F2) изучалась в отношении её роли в управлении движениями руки обезьяны с целью коснуться или схватить какой-то предмет . Нейроны в PMDc активировались в процессе движения руки с целью коснуться предмета. У обученной обезьяны нейроны в данной области были активны во время приготовления к хвату или касанию, а также в ходе самого хвата или касания. Наибольший отклик в данной области наблюдался при однонаправленном движении, несколько худший — при движениях в разные направления. Электрическая стимуляция области PMDc провоцировала комплексное движение плеча, руки, кисти, напоминающее то движение, которое возникает при приготовлении руки к хватанию .
Премоторная дорсальная ростральная область (PMDr, F7), вероятно, участвует в обучении, при котором произвольные сенсорные стимулы ассоциируются со специфическими движениями . Также данная область, вероятно, имеет отношение к движению глаз: электрическая стимуляция области PMDr вызывала движения глаз , активность нейронов в данной области соответствовала движениям глаз .
Премоторная вентральная каудальная область (PMVc, F4) изучалась в основном в отношении её роли в сенсорной регулировке движений ( англ. sensory guidance of movement). Нейроны в данной области чувствительны к тактильным, визуальным и слуховым стимулам . Эти нейроны особенно чувствительны к объектам, расположенным близко к телу, примерно на расстоянии вытянутой руки или в так называемом периперсональном пространстве ( англ. peripersonal space). Электрическая стимуляция данной области вызывает «защитные» движения, которые похожи оборонительную реакцию по «защите» тела . Эта премоторная область может являться частью более крупной системы, которая отслеживает безопасную дистанцию от объектов до тела и осуществляет в связи с этим оборонительные реакции .
Премоторная вентральная ростральная область (PMVr, F5) изучалась в основном в отношении её роли в работе мышц кисти руки при хватании и взаимодействии между рукой и ртом . Так, электрическая стимуляция нейронов в области F5 вызывала комплексное движение, при котором рука двигалась в сторону рта, кисть сжималась, сжатая кисть двигалась ко рту, шея поворачивалась, чтобы рот был на одной линии с поднесённой рукой, рот открывался .
Зеркальные нейроны были впервые обнаружены именно в зоне F5 в обезьяньем мозге в экспериментах Риццолатти и его коллег . Эти нейроны активировались, когда обезьяна хватала предмет, а также когда обезьяна наблюдала, как кто-то другой хватал предмет. Из-за этого предполагается, что нейроны в данной области являются как сенсорными, так и моторными. Зеркальные нейроны могут способствовать обучению путём имитации чужих действий и пониманию действий других особей.
В первых научных работах по моторным участкам коры исследователи выделяли только одну область, отвечающую за моторный контроль. Кэмпбелл в 1905 году стал первым исследователем, предположившим, что в коре может быть две моторные области, «первичная» и «промежуточная прецентральная». Свои предположения он обосновывал цитоархитектоникой. Первичная моторная кора содержит гигантские клетки Беца , которых очень мало или совсем нет в примыкающей коре.
В 1909 году Бродман на основе того же критерия выделил поле 4 (соответствующее первичной моторной коре) и поле 6 (соответствующее премоторной коре).
В 1919 году супруги Фогт ( Оскар и Сесиль) предположили, что моторная кора разделяется на первичную моторную кору (поле 4) и моторную кору более высокого порядка (поле 6), примыкающую к ней. Также, согласно их публикациям, поле 6 можно разделить на две части, 6a (дорсальную часть) и 6b (вентральную часть). Дорсальная часть дополнительно разделяется на область 6a-alpha (задняя часть, прилегающая к первичной моторной коре) и 6a-beta (передняя часть, прилегающая к префронтальной коре). Эти корковые области формируют иерархию, в которой область 6a-beta отвечает за контроль самых сложных движений, область 6a-alpha имеет «промежуточные свойства», а первичная моторная кора отвечает за самые простые движения. Таким образом, в работах Фогтов впервые появилась идея о выделении каудальной и ростральной премоторной коры.
В 1935 году Фултон подтвердил различия между первичной моторной областью в поле 4 и премоторной областью в поле 6. Свои выводы он основывал на изучении поражений соответствующих областей коры у обезьян. Неизвестно, кто первый использовал термин «премоторный», но именно Фултон популяризировал его.
И супруги Фогт, и Фултон отмечали, что различия в функциях премоторной и первичной моторной области не абсолютны, обе области напрямую связаны со спинным мозгом и действуют в какой-то мере «параллельно» друг другу, а не строго иерархично .
В 1937 году Пенфилд отрицал существование особой премоторной области. Он полагал, что нет никакого функционального отличия между полями 4 и 6. На его карте в задней части зоны 4 находились руки и пальцы, а в зоне 6 — мышцы спины и шеи.
В 1950-е годы Вулси , изучавший моторные карты обезьян, также не делал различий между первичной моторной и премоторной областями. Он использовал термин M1, которым обозначил области, включающие в себя премоторную и первичную моторную кору, и термин M2, которым обозначил медиальную моторную кору (в настоящее время эта область известна как дополнительная моторная область ).
После публикаций Пенфилда по человеческому мозгу и публикаций Вулси по обезьяньему мозгу к 1960-м годам идея о выделении латеральной премоторной коры отдельно от первичной моторной коры практически исчезла из научной литературы. Вместо этого выделялась область M1 как цельная структура, расположенная вдоль центральной борозды .
В 1980-е годы гипотеза об отдельной премоторной области вновь стала появляться в научных публикациях.
Роланд и коллеги изучали дорсальную премоторную кору и дополнительную моторную область человеческого мозга, проводя мониторинг кровотока в мозге посредством позитронно-эмиссионной томографии . Когда люди осуществляли сложные движения, основанные на сенсорных ориентирах (например, следовали вербальным инструкциям), усиливался кровоток в дорсальной премоторной коре. Когда люди осуществляли движения, ориентируясь на свой собственный «внутренний» темп, усиливался кровоток в дополнительной моторной области. Когда люди осуществляли простые движения, почти не требующие планирования, например, ощупывание предмета рукой, кровоток был в основном ограничен областью первичной моторной коры. Исходя из этих данных, первичная моторная кора активизируется во время простых движений, премоторная кора — во время движений, осуществляемых на основе поступающей сенсорной информации, а дополнительная моторная область — во время движений, осуществляемых на основе внутренних стимулов.
Уайз и коллеги изучали дорсальную премоторную кору обезьян. Обезьян учили выполнять задачу с задержанной ответной реакцией ( англ. delayed response task), в которой движение нужно было осуществить в ответ на определённый сенсорный сигнал. Во время выполнения задания нейроны в дорсальной премоторной коре активировались в ответ на сенсорный стимул и часто оставались активными в течение нескольких секунд задержки или приготовления, перед тем, как обезьяна выполняла нужное движение. Нейроны в первичной моторной коре были гораздо менее активными в фазе приготовления к движению и активизировались собственно в момент движения. Исходя из этих данных, дорсальная премоторная кора вовлечена в планирование и приготовление к движению, а первичная моторная кора — в осуществление движения.
Риццолатти и коллеги разделили премоторную кору на четыре части на основании цитоархитектоники: две дорсальные и две вентральные части. Затем они изучили свойства вентральных премоторных областей, включая тактильные, визуальные и моторные .
Было обнаружено, как минимум, три области, отвечающих за движение руки моторной коре: в первичной моторной, в вентральной премоторной и дорсальной премоторной коре . Исходя из этого можно предположить о существовании минимум трёх отличных корковых областей, каждая из которых выполняет собственную функцию в отношении движений пальцев и запястья.
После вышеуказанных публикаций возник новый консенсус, согласно которому существует не единый моторный корковый комплекс, как полагал Пенфилд, а несколько различных моторных областей, включающих первичную моторную и несколько премоторных участков, причём разные премоторные участки имеют отличные свойства и функции. Некоторые из них напрямую связаны со спинным мозгом и могут напрямую контролировать движения, другие же действуют опосредованно. Точная модель функционирования этих областей до сих пор остаётся неизученной.
Грациано и коллеги предложили альтернативную гипотезу организации первичной моторной коры и каудальной премоторной коры — кортикальных участков, напрямую связанных со спинным мозгом и включённым Пенфилдом и Вулси в моторную область M1. Согласно данной гипотезе, моторная кора организована в виде карты «естественного поведенческого репертуара» . Комплексная, многофакторная природа поведенческого репертуара объясняет комплексную, неоднородную карту коры, в которой различные части поведенческого репертуара отражаются в различных областях коры больших полушарий. Сложные движения, например, хватание предмета или лазание, требуют высокой координации разных частей тела, оценки местоположения близлежащих объектов, планирования на несколько секунд вперёд. Другие части репертуара движений, например, манипуляции с объектом при помощи пальцев, когда объект уже в руках, или манипуляции с объектом во рту, не требуют сложного планирования, просчёта траектории движения; здесь на первый план выходит контроль над движениями суставов и напряжением мелких мышц. Таким образом, более комплексные движения, особенно многосегментные, осуществляются при помощи передней части «моторной карты», так как эта часть коры связана с мышцами спины и шеи и координирует части тела. Более простые движения, связанные с дистальными мышцами, осуществляются более задними частями моторной коры. Согласно данной теории, хотя менее сложные движения связаны с первичной моторной корой, а более комплексные движения с каудальной премоторной корой, это не означает, что существует некая «иерархия» контроля движений. В каждой области есть свои «субрегионы» с отличными свойствами, что отражает неоднородность и комплексность репертуара движений.
Поражения премоторной коры могут вызывать апраксию — нарушение сложных форм произвольного целенаправленного действия при сохранности составляющих его элементарных движений, моторные персеверации — навязчивое воспроизведение одних и тех же движений или их элементов. У больных наблюдается затруднение воспроизведения предварительно заученных движений с использованием сенсорных ориентиров, при этом те же движения сравнительно легко заучиваются на основе пространственных ориентиров .