, где
x
— возраст, а
p
— относительная вероятность смерти за определённый промежуток времени,
a
и
b
— коэффициенты. Таким образом, при отсутствии постоянного члена
a
, размер популяции снижается с возрастом по двойной экспоненте
, где
x
— возраст, а
p
— относительная вероятность смерти за определённый промежуток времени,
a
и
b
— коэффициенты. Таким образом, при отсутствии постоянного члена
a
, размер популяции снижается с возрастом по двойной экспоненте
![s(x)=exp[-m(b^{x}-1)]](/images/000/140/140490/3.jpg?rand=406102) .
.

Старение человека
- 1 year ago
- 0
- 0

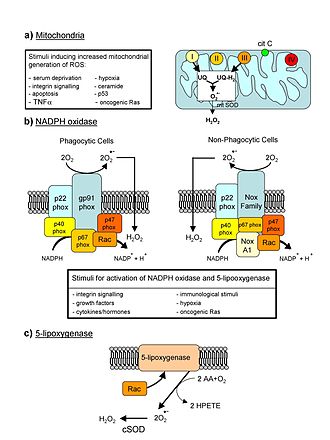
Популяционная динамика старения — направление исследования старения с помощью методов , то есть исследования возрастного состава популяций стареющих организмов и изменений этой зависимости в зависимости от типа организма и условий окружающей среды .
Наибольший интерес представляет динамика старения , включая человека , у которых старение наступает через большой промежуток времени после достижения половой зрелости и имеет постепенный характер. В отличие от одноплодных форм, многоплодным организмам не нужно использовать все жизненные силы до конца их воспроизводящей фазы (фазы размножения) для того, чтобы размножение прошло успешно, и средняя продолжительность жизни относительно периода размножения весьма значительно меняется у разных особей и в зависимости от вида: маленькие грызуны и дикие птицы используют в среднем только от 10 до 20 процентов их потенциального периода размножения, тогда как киты , слоны , обезьяны и другие крупные млекопитающие в природных условиях используют более 50 процентов своих периодов размножения, и часто даже переживают его.
Популяционный подход рассматривает зависимость размера популяции от возраста организмов. Изменения размера популяции с возрастом называются смертностью , что в случае стационарного состояния соответствует количеству организмов, которые умирают за единицу времени. Соответственно, относительные изменения размера популяции, или вероятность смерти за единицу времени, называются относительной смертностью. Обратным к смертности показателем, что также часто используется при описании популяционной динамики старения, есть вероятность выживания за единицу времени.
Целью популяционного подхода является определение закономерностей в зависимости размера популяции от времени, которые используются для определения скорости процесса старения. Эти данные, в свою очередь, могут использоваться для проверки моделей старения, выведенных или основываясь на физиологических и генетических механизмах, или с помощью общих системных механизмов.
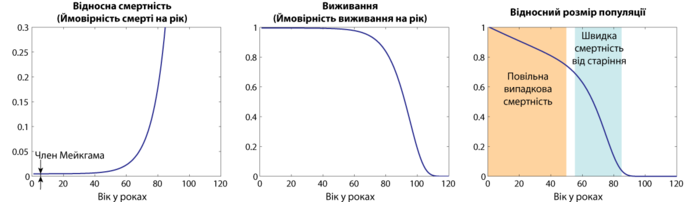
Величина, которая непосредственно измеряется — это возрастная зависимость популяции, из-за этого искомая величина является наиболее общей мерой смертности и старения. Тем не менее, более наглядной величиной является смертность или выживание — показатели, которые в большей мере характеризуют сам процесс старения. Часто используются логарифмические кривые, которые лучше отображают некоторые характерные черты приведенных зависимостей.
Одной из первых и наиболее распространённых сегодня математической моделью описания старения многоплодных организмов является так называемый закон смертности Гомпертца-Мейкхама (или просто Гомпертца), согласно которому вероятность смерти экспоненциально возрастает с возрастом: , где x — возраст, а p — относительная вероятность смерти за определённый промежуток времени, a и b — коэффициенты. Таким образом, при отсутствии постоянного члена a , размер популяции снижается с возрастом по двойной экспоненте .
Закон Гомпертца эмпирический и имеет место не для всех животных и не на всех промежутках времени, но он наиболее простой для сравнения старения разных организмов, и потому его коэффициенты часто используются в качестве показателей темпа (скорости) старения.

Коэффициент экспоненты функции Гомпертца показывает скорость старения. Различия в долголетии между видами является результатом прежде всего различий в скорости старения, и потому выражаются в различиях в этом коэффициенте.
Сравнение таблиц смертности разных линий мышей одного вида показывает, что различия между линиями в первую очередь происходят от различий в члене Мейкхама (независимому от возраста члене) функции Гомпертца. Если линии отличаются только независимым от возраста членом, менее долголетние линии имеют большую смертность, которая выше на постоянную величину на протяжении всей жизни, что проявляется в вертикальному сдвиге функции Гомпертца. При этом часто случается, что гибриды первого поколения (F1) двух естественных линий живут дольше, чем любой из родителей. Хотя исследований биохимических процессов таких гибридов не проводилось, таблицы продолжительности жизни указывают, что гибриды отличаются от родительских линий только независимым от возраста членом, но не сменой скорости старения. Другие исследования также показали, что в значительной мере вариации в продолжительности жизни между линиями мышей поясняются различиями в унаследованной склонности к определённым болезням.
Для популяций людей в разных странах к середине XX века разница в средней продолжительности жизни (не принимая во внимание детскую смертность ) практически исключительно происходила от разницы в члене Мейкхама. С середины XX века ситуация изменилась, что привело к почти параллельному переносу кривой смертности вправо. Хотя причины этого изменения неизвестны, предположительно они обусловлены существенным прогрессом личной и общественной гигиены , улучшением жилищных условий и медицинского обслуживания, качества питания, созданием эффективных вакцин и антибиотиков .
Закон Гомпертца-Мейкхама является только приближением, верным в среднем возрастном диапазоне. В области малого возраста наблюдается значительно более высокая смертность, чем предусматривается этим законом. Например, во время нереста может отложить до 6 миллионов икринок , но только небольшое количество из них доживает до момента половой зрелости . Эта смертность является преимущественно результатом неспособности молодёжи избегать хищников, бороться с болезнями, а также может быть результатом врождённых дефектов, и не является результатом старения.
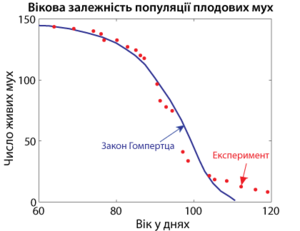
В области позднего возраста, наоборот, наблюдается уменьшение смертности по сравнению с законом Гомпертца, точнее выход вероятности смерти за единицу времени на плато . Как и в случае детской смертности, это общий закон, который наблюдается даже в неживой природе . И хотя одним из возможных пояснений этого явления могла бы быть гетерогенность популяции, современные данные чётко указывают на связь выхода смертности на плато с замедлением процесса старения .
Распространённым методом исследования старения является математическое моделирование популяционной динамики. Все математические модели старения можно приблизительно разбить на два главных типа: модели данных и системные модели . Модели данных или аналитические модели — это модели, которые не используют и не пытаются пояснить какие-либо гипотезы о физических процессах в системах, для которых эти данные получены. К моделям данных относятся, в частности, и все модели математической статистики. В отличие от них, системные или механистические модели строятся преимущественно на базе физических законов и гипотез о структуре системы, главным в них является проверка предложенного механизма.
Ниже приведен список важнейших из предложенных математических моделей :
Первыми из этих математических моделей начали появляться модели данных смертности. Ещё задолго до понимания и даже исследования процессов, которые лежат в основе старения, возник практический интерес к прогнозу будущей продолжительности жизни для использования в страховании и демографии . Именно для расчёта страховых премий ещё в XIX веке были разработаны первые таблицы смертности и сформулированы известные модели Гомпертца и Гомпертца-Мейкхама . Введя простую двухпараметрическую модель смертности, Гомпертц дал исследователям возможность не только рассчитывать будущие шансы долголетия, но и исследовать изменения двух фундаментальных параметров: начальной смертности и темпа старения. Выделение из данных смертности возрастной компоненты дало начало математической геронтологии .
В конце XX века начало возникать много новых демографических моделей смертности. Получение значительного количества новых данных, часто для гетерогенных популяций, привели к новым методам анализа таблиц смертности . Эти модели, часто с использованием методов анализа стохастических процессов, позволили выделение отдельных компонентов смертности и описание влияния заболеваний и факторов окружающей среды на долголетие .
Из-за того, что уже давно было известно о линейности снижения функциональных возможностей организма , возникла необходимость увязки этой динамики с экспоненциальным возрастанием с возрастом относительной смертности. Одним из первых объяснений этого явления стала модель Стрелера-Милдвана . Эта модель предполагает флуктуации системы, вероятность которых экспоненциально падает с размером. Для их преодоления организм должен затрачивать энергию, но максимальные затраты линейно снижаются с возрастом. В результате вероятность смерти, то есть невозможность преодолеть флуктуацию, экспоненциально возрастает. Похожей моделью является модель Сечера-Трукко , которая предлагает гауссовское распределение внешнего влияния, и с некоторыми условиями также приводит к экспоненциальной зависимости смертности от возраста. Альтернативным подходом к пояснению этой зависимости является использование теории надёжности , которая поясняет экспоненциальную зависимость через значительную чрезмерность биологических систем .
Следующим шагом в математическом моделировании стало пояснение отклонения смертности от классической зависимости, в первую очередь выхода смертности на плато в позднем возрасте. В целом предложенные модели являются модификациями упомянутой модели Стрелера-Милдвана с использованием стохастических данных, наиболее известной является модель Мюллера и Роуза . Кроме этой модели было предложено несколько модификаций, например, модификация, основанная на теории надёжности . Стохастический подход, который лежит в основе, предлагает уменьшение эффекта чрезмерности биологической системы за счёт выделения наиболее занятых каналов повреждения системы . Ещё один подход, основанный на гетерогенности популяции, показал неспособность пояснить экспериментальные данные . Эволюционный подход к проблеме выхода относительной смертности является модификацией модели Гамильнона , основанной на принципе антагонистической плейотропии . Идея состоит в том, что давление естественного отбора снижается для мутаций, которые связаны с изменениями, которые проявляются только в позднем возрасте, но не обязательно достигает нуля , например за счёт эффектов, связанных с увеличением ценности опытных старых организмов по сравнению с молодыми, несмотря на уменьшение их числа .
Системные модели в целом рассматривают много отдельных факторов, событий и явлений, которые непосредственно влияют на выживание организмов и рождения потомства. Эти модели, основываясь на теории одноразовой сомы , в целом рассматривают старение как баланс и перераспределение ресурсов как в физиологическом (в течение жизни одного организма), так и в эволюционном аспектах. Как правило, особенно в последнем случае, речь идёт о распределении ресурсов между непосредственными затратами на рождение потомства и затратами на выживание родителей . Многие из этих моделей, список которых приведен выше, основываются на методах статистического моделирования. Часто рассматривается вопрос о адекватности моделей истории жизни результатам экспериментов на животных, в первую очередь популяционным данным.