Дейноних
- 1 year ago
- 0
- 0

Дейноних ( лат. Deinonychus — от др.-греч. δεινός/deinós — «ужасный» и ὄνυχος/ónychos ( р. п. от др.-греч. ὄνυξ/ónуx) — «коготь») — род хищных тероподовых динозавров из семейства дромеозаврид , в который включают единственный вид — Deinonychus antirrhopus . Ископаемые остатки найдены в нижнемеловых отложениях (115—108 миллионов лет назад) на территории США — в штатах Монтана , Юта , Вайоминг и Оклахома , хотя зубы, которые могут принадлежать дейнониху, обнаружены также в штате Мэриленд .
Исследования палеонтолога Джона Острома , посвящённые дейнонихам в конце 1960-х — начале 1970-х годов произвели революцию в понимании учёными динозавров, что привело к « » и вызвало споры о том, были ли динозавры теплокровными или холоднокровными. До этого динозавры виделись учёным как огромные, медлительные и несообразительные животные. Остром отметил небольшое тело, горизонтальную осанку, позвоночник, похожий на страусиный, и особенно увеличенные серповидные когти на ногах, что вместе указывало на активного, ловкого хищника.
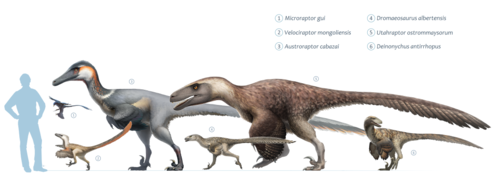
Дейноних был некрупным динозавром размером до 3,3 метра и оценочной массой от 73 до 100 килограммов, узкой мордой и широкими скулами, дававшими ему возможность стереоскопического зрения. Несмотря на отсутствие связанных с дейнонихом отпечатков мягких тканей, имеются свидетельства о наличии у дромеозаврид перьев. Скорее всего, дейноних также имел развитый перьевой покров.
Как в формации Кловер, так и в формации Энтлерс остатки дейнонихов часто находят вместе с остатками птицетазовых динозавров тенонтозавров , на которых дейнонихи охотились или, по меньшей мере, объедали их останки.

Окаменелые остатки дейнониха были обнаружены в геологической формации , расположенной в штатах Монтана и Вайоминг , и в современной ей формации , штат Оклахома . Формацию Кловерли датируют поздней аптской — ранней альбской эпохами раннего мелового периода (115—108 миллионов лет) . Кроме того, к роду могут быть отнесены зубы, найденные в штате Мэрилэнд — в формации (среднеаптская эпоха) , расположенной в Приатлантической низменности .
Первые окаменелости дейнониха были обнаружены в 1931 году в южной части штата Монтана недалеко от города Биллингс . Руководитель группы, палеонтолог Барнум Браун , прежде всего занимался раскопками и препарацией остатков динозавра- орнитопода тенонтозавра , но в своём полевом отчёте с места раскопок для Американского музея естественной истории он сообщил об обнаружении рядом со скелетом тенонтозавра остатков небольшого плотоядного динозавра, «но заключённого в известняк и трудно поддающегося препарации» . Учёный неофициально назвал новое животное «Daptosaurus agilis» , собираясь описать его и выставить напоказ скелет, но так и не закончил работу. Браун привёз из формации Кловерли скелет теропода меньшего размера, но с более крупными зубами, которого он назвал «Megadontosaurus» . Спустя десятилетия, изучавший этот материал Джон Остром понял, что зубы принадлежали дейнониху, а скелет — совершенно другому животному. Остром назвал этого динозавра микровенатор .

Чуть более 30 лет спустя после раскопок Брауна, в августе 1964 года, Джон Остром возглавил экспедицию Йельского университета , которая обнаружила обширный скелетный материал около города , штат Монтана. В течение следующих двух лет участники экспедиций раскопали более 1000 костей, принадлежавшие по меньшей мере трём особям. Кости были найдены несочленёнными, что делало невозможным определение точного числа отдельных представленных животных, поэтому типовой образец дейнониха (YPM 5205) был ограничен полной левой и частичной правой стопами , которые определённо принадлежали одной особи. Остальные кости были внесены в каталог музея под 50 отдельными каталожными индексами, несмотря на то, что принадлежали нескольким животным .
В более позднем исследовании Джон Остром и Грант Мейер проанализировали собранный материал, а также остатки «Daptosaurus» Барнума Брауна, и обнаружили, что образцы принадлежат одному и тому же виду. Остром впервые опубликовал свои выводы в феврале 1969 года, дав всем упомянутым окаменелым остаткам новое имя Deinonychus antirrhopus . Название рода происходит от др.-греч. δεινός/deinós — «ужасный» и ὄνυχος/ónychos ( р. п. от др.-греч. ὄνυξ/ónуx) — «коготь»). Видовое название лат. antirrhopus происходит от др.-греч. ἀντίρροπος и переводится как «уравновешенный» и относится к вероятной цели укреплённого связками хвоста . В июле 1969 года Остром опубликовал обширную монографию, посвящённую дейнониху .
Хотя к 1969 году было известно множество костей дейнониха, многие важные из них было трудно интерпретировать или они вовсе отсутствовали. Были известны некоторые заглазничные элементы черепа, но не были найдены бедренные кости , крестец , вилочка или грудина , отсутствовали множество позвонков и (как считал Остром) имелся только крошечный фрагмент коракоида . Выполненная Остромом скелетная реконструкция дейнониха включала очень необычную часть таза — лобковую кость , которая была трапециевидной и уплощённой, в отличие от таковой у других теропод, но была такой же длины, что и седалищная кость , и была найдена прямо рядом с ней .

В 1974 году Остром опубликовал ещё одну монографию о плечевом поясе дейнониха, в которой он признал, что лобковая кость, описанная ранее, на самом деле является коракоидом — элементом плеча . В том же году в Монтане члены экспедиции Гарвардского университета нашли и откопали ещё один образец дейнониха (MCZ 4371). Это открытие добавило несколько новых элементов скелета: хорошо сохранившиеся бедренные и лобковые кости, крестец и лучше сохранившуюся подвздошную кость, а также элементы стоп и плюсны . Остром описал этот образец и внёс исправления в остеологию динозавра. На этот раз скелет имел очень длинный лобок, и Остром стал подозревать, что эта кость была несильно загнутой назад, совсем как у птиц .

Но даже после всех статей, написанных Остромом, и работы, которая им предшествовала, в Американском музее естественной истории оставалось несколько небольших блоков с ископаемым материалом. Они состояли в основном из изолированных костей и костных фрагментов, заключённых в исходную матрицу из скальных пород, в которой первоначально были депонированы образцы. В 2000 году эти образцы изучили Жерар Грелле-Тиннер и Питер Маковицки. Они выявили одну интересную особенность, упущенную из виду ранее. Оказалось, что несколько длинных тонких костей, идентифицированных на блоках как окостеневшие сухожилия (структуры, которые помогали придать жёсткость хвосту дейнониха), на самом деле являлись гастралиями — брюшными рёбрами. Что ещё более важно, в оригинальной каменной матрице, которая окружала образец животного, было обнаружено большое количество яичной скорлупы .
В последующем, более подробном отчёте о яичной скорлупе Грелле-Тиннер и Маковицки пришли к выводу, что яйцо почти наверняка принадлежало дейнониху и представляло первое идентифицированное яйцо дромеозаврид . Кроме того, внешняя поверхность одной яичной скорлупы была обнаружена в тесном контакте с гастралией, что позволяет предположить, что этот динозавр мог насиживать яйца. Это, в свою очередь, подразумевало использование животным теплообмен тела в качестве механизма для инкубации яиц, и указывает на эндотермию , как у современных птиц . Дальнейшее исследование, проведённое группой учёных под руководством , показало, что животному на момент смерти было 13 или 14 лет и его размер был максимально возможным. В отличие от других теропод, обнаруженных вместе с яйцами и гнёздами, этот динозавр был полностью выросшим .
Описание дейнониха, выполненное Джоном Остромом в 1969 году, признано единственным важным открытием в палеонтологии динозавров середины XX века . Открытие этого явно активного, ловкого хищника во многом изменило научную (и популярную) концепцию динозавров и дало возможность предположить, что некоторые динозавры могли быть теплокровными. Эту концепцию назвали . Несколько лет спустя Остром отметил сходство между передними лапами дейнониха и птиц: это наблюдение привело его к возрождению гипотезы о происхождении птиц от динозавров .
Опираясь на исследования анатомии, чрезвычайно похожей на птичью, а также на сильную связь с другими дромеозавридами, многие палеонтологи выдвинули гипотезу, согласно которой дейноних был покрыт перьями . Явные ископаемые свидетельства о наличии перьевого покрова существуют для нескольких родственных дромеозаврид, включая велоцираптора и микрораптора , хотя прямых доказательств для самого дейнониха пока не известно . В исследованиях, посвящённых диапазону движений передних конечностей дромеозаврид, учитывалось наличие у них маховых перьев, которые присутствуют у всех образцов дромеозаврид с отпечатками мягких тканей .
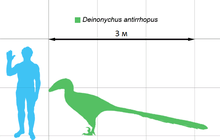
Основываясь на нескольких экземплярах зрелых особей , мы знаем, что дейнонихи могли достигать 3,3 метра в длину с длиной черепа в 410 миллиметров, высоты в бедре 0,87 метра и массы в 73 килограмма , хотя имеются и более высокие оценки массы — до 100 килограммов . Череп нёс мощные челюсти, оснащённые 70 загнутыми, лезвиеподобными зубами. За десятилетия, прошедшие со времени первых находок, исследования черепа значительно продвинулись. Остром восстановил неполные, плохо сохранившиеся черепа дейнонихов как треугольные, широкие и очень похожие на черепа аллозавров . Дополнительный черепной материал дейнонихов и близкородственных видов, обнаруженный в хорошем состоянии в трёхмерном виде , показал, что нёбо у этого динозавра было более сводчатым, чем думал Остром, что делало морду животного намного более узкой, в то время, как скуловые кости были широкими, что давало дейнониху возможность стереоскопического зрения . При этом череп дейнониха отличался от черепа велоцираптора наличием более прочной крышки черепа, как у дромеозавра , и отсутствием вдавленных и уплощённых ноздрей велоцираптора . И череп, и нижняя челюсть содержали отверстия , уменьшавшие вес костей. Предглазничное отверстие у дейнониха было особенно большим .
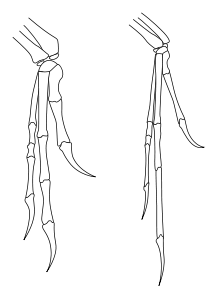
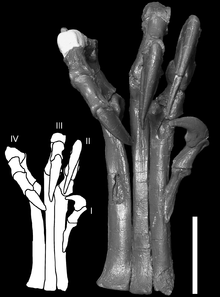
У дейнониха были большие кисти с тремя когтями на каждой конечности. Первый палец был самым коротким, второй — самым длинным. Второй палец на задней конечности нёс крупный серповидный коготь, который, возможно, использовался во время охоты .
Рядом с фоссилиями дейнониха никогда не находили отпечатков кожи. Тем не менее, данные свидетельствуют, что у дромеозаврид, включая дейнониха, были перья . Многочисленный окаменелости микрораптора, представителя того же семейства, более древнего геологически и более примитивного филогенетически , тем не менее сохранили отпечатки перьевого покрова, аналогичного таковому у современных птиц . Велоцираптор геологически моложе дейнониха, но тесно с ним связан. На локтевых костях этого динозавра обнаружили перьевые бугорки — места крепления маховых перьев; перьевые бугорки являются прямым индикатором наличия перьев современного типа .
Положение дейнониха в систематике постоянно меняется внутри семейства дромеозаврид, в связи с регулярным открытием новых представителей группы и проводимых авторами описаний филогенетических анализов . Сам Джон Остром начиная с 1969 года помещал открытого им динозавра в семейство дромеозаврид ; туда же дейнониха относило большинство исследователей . Другие авторы долгое время сближали дейнониха с велоцираптором, помещая оба таксона в подсемейство велоцирапторин . Филогенетическое исследование 2020 года, проведённое командой палеонтологов под руководством Стивена Ясински при описании вида Dineobellator notohesperus , поместило дейнониха в группу Eudromaeosauria , вне какого-либо подсемейства .
| Eudromaeosauria |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма 2021 года :
| Dromaeosauridae |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В исследовании 2022 года род Deinonychus реклассифицировали как базальный в кладе Dromaeosaurinae .

В формации Кловерли очень часто рядом с остатками растительноядного динозавра тенонтозавра находят зубы дейнониха. В двух карьерах рядом с окаменелыми остатками тенонтозавров сохранились довольно полные скелеты дейнонихов. Первый, Йельский карьер, на территории штата Монтана, содержит остатки 4 взрослых и 1 неполовозрелой особей дейнонихов, а также многочисленные зубы этих животных. Присутствие такого количества скелетов дейнонихов в одном карьере предполагает, что дейнонихи могли питаться тенонтозавром и, возможно, охотиться на него. В 1995 году Остром и Максвелл использовали эту находку, чтобы заявить, что дейнонихи охотились в стаях . Второй такой карьер находится в формации Энтлерс, штат Оклахома. Здесь обнаружили 6 неполных скелетов тенонтизавров разных размеров, а также один частичный скелет и множество зубов дейнониха. Одна плечевая кость тенонтозавра имеет следы, которые интерпретируют как царапины зубов дейнониха. В 1998 году группа учёных под руководством Д. Бринкмана отметили, что масса взрослого дейнониха равнялась 70—100 килограммов, а масса тенонтозавра — от 1 до 4 тонн. В одиночку дейноних не смог бы справиться с тенонтозавром, что, по мнению учёных, указывает на стайное поведение хищника .
Проведённое в 2007 году исследование Д. Бринкмана и Б. Роуча, основанное на знаниях об охоте современных хищников и тафономии тенонтозавра, поставило под сомнение стайный способ охоты дейнониха. Современные архозавры , такие, как птицы и крокодилы , а также комодские вараны обычно не охотятся в стаях — они либо охотятся в одиночку, либо объедают туши уже мёртвых животных, когда и происходят конфликты между особями одного вида. Например, в ситуациях, когда комодские вараны совместно поедают тушу, сначала едят более крупные особи. Они атакуют мелких варанов, если те пытаются кормиться; если меньшее животное погибает, сородичи его съедают. Фоссилии, найденные в вышеописанных карьерах, указывают на стратегию питания дейнонихов, очень похожую на таковую у комодских варанов или крокодилов. Самые неполные скелеты здесь принадлежат неполовозрелым особям, а недостающие части могут указывать на то, что их съели другие дейнонихи . С другой стороны, найдены цепочки следов параллельно идущих животных с равным расстоянием между шагами, что подразумевает стайный образ жизни . Также подмечено, что крокодилы всё-таки охотятся совместно , что ставит под сомнение гипотезу о борьбе за пищу и каннибализм среди дейнонихов.
В 2011 году Денвер Фаулер и его коллеги предложили новый метод, согласно которому дейноних и другие дромеозавриды могли захватывать и удерживать добычу . Эта модель, известная как «хищник удерживает жертву», предполагает, что дейноних убивал свою жертву тем же способом, каким её убивают современные хищные птицы: прыгнув на свою добычу, динозавр придавливал её своим весом и крепко удерживал своими серповидными когтями. Как и птицы, дромеозавриды поедали свою добычу живой. Это предложение основано, главным образом, на сравнении морфологии и пропорций стоп и голеней дромеозаврид с несколькими группами существующих хищных птиц с известным хищническим поведением. Фаулер обнаружил, что ступни и ноги у дромеозаврид наиболее похожи на ноги орлов и ястребов , особенно в том, что касается увеличенного второго когтя и схожего диапазона хватательных движений. Однако, короткая плюсневая кость и сила стопы были бы больше похожи на таковые у сов. Этот метод хищного поведения коррелирует с другими аспектами анатомии дейнониха, такими, как необычная морфология челюстей и кистей. Передние конечности, вероятно, были покрыты длинными перьями, и, возможно, использовались в качестве стабилизаторов колебания для равновесия во время борьбы с добычей, наряду с жёстким уравновешивающим хвостом. Его челюсти, которые, как считается, имели сравнительно слабую силу укуса , могли производить движения, подобные движениям пилы. Такие пилящие движения производят челюсти комодского варана, который также имеет слабую силу укуса, чтобы прикончить свою добычу, если его атака не была достаточно мощной .

Впервые оценки силы укуса дейнониха была получена в 2005 году после реконструкции мускулатуры челюстей. Исследователи пришли к выводу, что этот динозавр, вероятно, имел максимальную силу укуса лишь 15 % от силы современного миссисипского аллигатора . В ходе исследования 2010 года, проведённого Полем Жиньяком и его коллегами, была сделана попытка оценить силу укуса, основываясь непосредственно на недавно обнаруженных следах проколов зубов дейнониха на костях тенонтозавра. Эти проколы были сделаны взрослой особью и явились доказательством того, что крупный дейноних мог прокусить кость. Команда Жиньяка смогла определить, что сила укуса дейнониха была значительно выше, чем её указали в ранних работах, и которая оценивалась только с помощью биомеханических исследований. Они обнаружили, что сила укуса этого динозавра составляет от 4100 до 8200 ньютонов, что больше, чем у живых хищных млекопитающих, включая гиену , и эквивалентна укусу аллигатора такого же размера .
При этом Жиньяк и его команда отметили, что следы проколов зубов дейнониха являются относительно редкой находкой, и, в отличие от более крупных теропод, таких, как тираннозавры , дейнонихи, вероятно, не часто кусали и грызли кости. Вместо этого они, вероятно, использовали свою высокую силу укуса для защиты или захвата добычи, а не для кормления .

Несмотря на то, что серповидный коготь является наиболее отличительной чертой анатомии дейнониха, его форма и кривизна варьируется между образцами. Типовой образец, описанный Остромом в 1969 году, имеет сильно изогнутый серповидный коготь, в то время как образец, описанный в 1976 году, имеет коготь с гораздо меньшей кривизной, более похожий по профилю на другие когти на оставшихся пальцах ног. Остром предположил, что различие в размере и форме серповидных когтей может быть связано с индивидуальными, половыми или возрастными вариациями, но признал, что не берётся утверждать с уверенностью .
Остром предположил, что дейноних орудовал своим когтем, чтобы резать и вспарывать кожу своих жертв . Некоторые учёные были уверены, что дейноних потрошил цератопсовых динозавров при помощи когтя . Другие исследования показали, что динозавр мог наносить только небольшие проколы . В 2005 году Фил Мэннинг и его коллеги провели испытания роботизированной копии, которая точно соответствовала анатомии дейнониха и велоцираптора, и была оснащена гидравлическими цилиндрами, чтобы заставить робота поразить тушу свиньи. Во время этих испытаний когти сделали только мелкие проколы и не смогли порезать или вспороть тушу. Авторы предположили, что когти были бы более эффективными при лазании, чем при нанесении смертельных ударов .
Остром сравнил дейнониха со страусом и казуаром . Он отметил, что эти птицы могут нанести серьёзные травмы большим когтем на втором пальце ноги . У казуара есть когти длиной до 125 миллиметров . Остром указал, что казуары могут разорвать руку или вспороть кожу человека . В 1999 и 2003 годах Кристофер Кофрон изучил 241 задокументированное нападение казауров и обнаружил, что, хотя один человек и две собаки были убиты, нет никаких случаев потрошения или расчленения этими птицами кого-либо . Казуары используют свои когти, чтобы защитить себя или дать отпор побеспокоившим их животным, при этом они предпочитают скорее демонстрировать угрозу, чем проявлять её . Кариамовые также имеют увеличенный коготь на втором пальце ноги и используют его, чтобы разорвать добычу на мелкие куски, которые можно проглотить . Исследование 2011 года показало, что серповидный коготь, скорее всего, использовался для того, чтобы придавить добычу во время укуса, а не в качестве режущего оружия .
Существуют анатомические доказательства, подтверждённые отпечатками следов, что серповидный коготь удерживался над землёй, пока динозавр шёл, наступая на третий и четвёртый пальцы ноги .

Биомеханические исследования, проведённые в 2002 году Кеном Карпентером , подтвердили, что наиболее вероятная функция передних конечностей заключалась в хватании, поскольку передние лапы дейнониха были пропорционально длиннее лап других теропод. Довольно большой вытянутый коракоид , указывающий на мощные мышцы передних конечностей, говорил в пользу этой интерпретации. Те же исследования показали, что дейноних не мог складывать лапы-крылья вдоль тела, по-птичьи , как это было указано в ранних работах Жака Готье и .
Исследования, проведённые Филом Сентером в 2006 году, показали, что дейноних мог использовать передние конечности не только для хватания, но и для прижимания предметов в направлении груди. Если бы у этого динозавра были оперённые пальцы и крылья, перья до некоторой степени ограничивали бы диапазон движения передних конечностей. Например, когда дейноних вытягивал конечность вперёд, кисть автоматически поворачивалась в положение вверх. Это привело бы к тому, что одно крыло заблокировало бы другое, если бы обе передние конечности вытягивались одновременно, что привело бы к тому, что прижимание предметов к грудной клетке могло бы осуществляться только одной конечностью за раз. Функция пальцев также была бы ограничена перьями; например, только третий палец передней лапы мог использоваться в таких действиях, как ощупывание мелких предметов, и только в положении, перпендикулярном крылу . Тем не менее, годом ранее, исследовании механики передних конечностей дейнониха Алан Гишлик обнаружил, что даже если бы присутствовали большие контурные перья, хватательная способность конечности не была бы значительно затруднена; скорее, динозавр осуществлял схватывание перпендикулярно крылу, и вероятно, удерживал бы объекты обеими лапами одновременно, как в «медвежьих объятиях» .
Уильям Парсонс и Кристин Парсонс показали, что неполовозрелые особи дейнонихов демонстрируют некоторые морфологические отличия от взрослых. Например, передние конечности молодых особей были пропорционально длиннее, чем у взрослых, что является возможным признаком различий в поведении между молодыми и взрослыми особями . Другим примером может служить функция серповидного когтя на ногах. Парсонсы предположили, что кривизна когтя (на разницу которой в 1976 году указывал ещё Остром ), вероятно, была выражена сильнее у ювенильных особей, поскольку это помогало им лазать по деревьям, а когти становились более выпуклыми, когда животное становилось старше и начинало жить исключительно на земле . Этот вывод был основан на гипотезе, что некоторые маленькие дромеозавриды использовали свои серповидные когти для лазания .
В 2015 году Уильям и Кристин Парсонсы провели исследование образца MCZ 8791 — дейнониха в возрасте от 1 до 2 лет. Учёные пришли к выводу, что приблизительный размер тела, длина передних конечностей, развитие запястья и наличие более заметного локтевого отростка на локтевой кости MCZ 8791 подтверждают гипотезу о том, что молодые особи этого таксона обладали некоторой способностью летать .

В популярных источниках дромеозаврид, особенно дейнонихов, часто изображают как быстро бегающих животных, и сам Джон Остром в первоописании предположил, что этот динозавр был быстрым животным . Однако, когда Остром описывал дейнониха, полных задних конечностей известно не было, и предположение Острома о длине бедренной кости оказалось завышенным. В более позднем исследовании учёный отметил, что отношение бедренной кости к большеберцовой кости не так важно при определении скорости, как относительная длина стопы к голени. У современных быстроногих птиц, таких как страусы, соотношение стопы к голени составляет 0,95. У быстро бегавших динозавров, таких как струтиомим , это соотношение составляет 0,68, но у дейнониха соотношение равняется 0,48. Остром заявил, что «единственный разумный вывод» заключается в том, что дейноних, хотя и далеко не медленный, не был особенно быстр по сравнению с другими динозаврами и, конечно, не так быстр, как современные бескилевые птицы .
Отношение длины ступни к длине голени у дейнониха частично связано с необычно короткими плюсневыми костями , и это соотношение было больше у молодых особей. Остром предположил, что короткая плюсна может быть связана с функцией серповидного когтя: по-видимому, коготь становится тем короче, чем старше становилось животное. Он истолковал все эти черты (короткий второй палец ноги с увеличенным когтем, короткую плюсну и т. д.) как доводы в пользу использования задней ноги в качестве наступательного оружия, когда серповидный коготь наносил удары одновременно вниз и назад, пока динозавр старался ранить жертву. Остром предположил, что короткая плюсна уменьшала общее напряжение костей ног во время такой атаки, и интерпретировал необычное расположение мышечных сухожилий в ноге дейнониха таким образом, что при нападении на добычу работали иные мышцы, чем при ходьбе или беге. Поэтому Остром пришел к выводу, что ноги дейнониха представляют собой баланс между адаптациями к бегу, необходимыми для ловкого хищника, и функциями, снижающими напряжение в костях, чтобы компенсировать своё уникальное ножное оружие .
В своём исследовании следов канадских динозавров, проведённом в 1981 году, палеонтолог Ричард Кул дал приблизительные оценки скорости движения, основанные на нескольких цепочках следов, оставленных разными видами в формации в Британской Колумбии . По оценкам Кула, животное, оставившее одну из этих цепочек, представляющую ихнотаксон Irenichnites gracilis и которая, возможно, была оставлена дейнонихом, имело скорость движения 10,1 км в час .

Идентификация в 2000 году вероятного яйца дейнониха, связанного с одним из оригинальных экземпляров, позволила сравнить его с яйцами других тероподовых динозавров, с точки зрения структуры яйца, а также привычек гнездования и размножения. При исследовании образца в 2006 году Ж. Грелле-Тиннер и П. Маковицки изучили возможность того, что дромеозаврид питался яйцом или что фрагменты яйца были связаны со скелетом дейнониха по совпадению. Они опровергли идею о том, что яйцо было пищей для животного, отметив, что фрагменты были зажаты между гастралиями и костями передних конечностей, что сделало невозможным их нахождение среди содержимого желудка животного. Кроме того, способ, которым яйцо было раздроблено и фрагментировано, указывало, что оно было целым во время захоронения и сломалось только в процессе окаменения. Идея, что яйцо было случайно связано с динозавром, также оказалась маловероятной; кости, окружающие яйцо, не были разбросаны или вычленены, а оставались нетронутыми относительно их положения при жизни, что указывает на то, что область вокруг яйца, включая само яйцо, не была нарушена во время консервации. Тот факт, что эти кости были гастралиями, которые очень редко бывают сочленены, подтверждает эту интерпретацию. Все свидетельства, согласно авторам исследования, указывают на то, что яйцо лежало неповреждённым под телом дейнониха, когда оно было похоронено. Вполне возможно, что данная находка представляет собой гнездовое поведение у дейнониха, сходное с тем, которое наблюдается у родственных троодонтид и овирапторид , или что яйцо фактически находилось внутри яйцевода , когда животное погибло .
Изучение микроструктуры яйца подтверждает, что оно принадлежало тероподу, поскольку имеет общие характеристики с другими известными яйцами теропод и показывает различия с яйцами птицетазовых и завропод . По сравнению с яйцами других манирапторных теропод яйцо дейнониха больше всего похоже на яйца овирапторид, чем троодонтид, несмотря на исследования, которые показывают, что последние более тесно связаны с дромеозавридами, такими как дейноних. Найденное яйцо было раздавлено, что не позволило определить его размер; Грелле-Тиннер и Маковицки оценили его диаметр приблизительно в 7 сантиметров, основываясь ни ширину тазового канала, через которое оно должно было пройти. Размер схож с диаметром самого крупного яйца овирапторида Citipati — 7,2 сантиметра; Citipati и дейноних также имели одинаковый общий размер тела, что подтверждает эту оценку. Кроме того, толщина яичной скорлупы Citipati и дейнониха практически одинакова, и поскольку толщина скорлупы коррелирует с объёмом яйца, это ещё раз подтверждает идею о том, что яйца этих двух животных были примерно одинакового размера .
Исследование, которое опубликовали в ноябре 2018 года М. Норелл, Т. Янг и Я. Виман, указывает на то, что дейнонихи откладывали яйца синего цвета, вероятно, чтобы замаскировать их в построенных ими открытых гнёздах. Исследование также указывает, что дейнонихи и другие динозавры, которые строили открытые гнёзда, вероятно, являются основателями происхождения цвета в современных птичьих яйцах в качестве адаптации как для распознавания, так и для маскировки. Авторы отметили, что их наличие цветных яиц уже более столетия считалось уникальной чертой для птиц. Однако, как оказалось, развитие маскировочной окраски яиц у динозавров шло одновременно с привычкой строить открытые гнёзда .
Геологические данные свидетельствуют о том, что дейноних обитал в пойменной или болотистой среде обитания . Древняя окружающая среда как формации Кловер, так и формации Энтлерс представляла собой тропические или субтропические леса, дельты и лагуны , мало чем отличающихся от экосистемы современной Луизианы . Другие животные, с которыми дейноних делил среду обитания, включают анкилозавра Sauropelta , орнитопод Zephyrosaurus и Tenontosaurus . На территории Оклахомы также были найдены остатки крупного теропода акрокантозавра , гигантского завропода завропосейдона , крокодиломорфов и , а также остатки панцирниковых щук . Если зубы, найденные в Мэриленде, принадлежали дейнониху, то его соседями были завропод Astrodon и известный только по окаменевшим зубам Priconodon .

Дейноних был описан в романах Майкла Крайтона « Парк юрского периода » и « », а также в серии одноимённых фильмов . Тем не менее, Крайтон в конечном итоге решил использовать для этих динозавров название « велоцираптор » вместо «дейнониха». В процессе написания романа писатель несколько раз встречался с Джоном Остромом, чтобы обсудить детали возможных моделей поведения и внешнего вида дейнониха. Крайтон в какой-то момент виновато сказал Острому, что решил использовать название «велоцираптор» вместо дейнониха для своей книги, поскольку прежнее имя было «более драматичным». Несмотря на это, по словам Острома, Крайтон заявлял, что велоцираптор в романе был основан на дейнонихе почти во всех деталях, и что он изменил только название животного .
Создатели фильма «Парк юрского периода» последовали его примеру, разработав модели, основанные почти полностью на дейнонихе, а не на велоцирапторе, и, как сообщается, во время производства они запросили все опубликованные работы Острома о дейнонихе . В результате в фильме они изобразили динозавров с размером, пропорциями и формой морды дейнониха .