
Орнитомим
- 1 year ago
- 0
- 0

Орнитоми́м ( лат. Оrnithomimus ; от др.-греч. ὄρνις — птица и μῖμος — подражание, воспроизведение; птичий имитатор) — род всеядных двуногих динозавров семейства орнитомимидов , живший в позднем меловом периоде на территории современной Северной Америки . На данный момент к роду принадлежат только два вида: типовой вид Ornithomimus velox и Ornithomimus edmontonicus . Семнадцать других наименований видов были отнесены к собственным родам или оказались непричастными к Ornithomimus velox .

Орнитомимы были быстрыми двуногими тероподами , чьи ископаемые свидетельства показывают наличие перьевого покрова. Подобно другим представителям своего семейства, орнитомимы характеризуются трёхпалыми опорными ступнями, длинными тонкими передними конечностями и длинными шеями с по-птичьи удлинёнными беззубыми клювами. Внешне они сильно напоминали страусов . Их ноги были сильными и длинными, кости — полыми, глаза и мозг — большими. Мозг орнитомимид вообще был большим для динозавров, но это не обязательно являлось признаком высокого интеллекта; некоторые палеонтологи считают, что увеличенные участки мозга отвечали за координацию движений . Кости лап по внешнему виду удивительно напоминают лапы ленивца , что натолкнуло Генри Осборна к мысли об использовании динозавром передних лап для притягивания веток с листвой во время кормления.
Орнитомимы отличаются от прочих представителей своего семейства, таких, как струтиомим , более короткими торсами, длинными тонкими предплечьями, очень тонкими прямыми когтями на кистях и стопах, а также одинаковой длиной пястных костей и фаланг пальцев .
Два вида орнитомимов сегодня рассматриваются как по-настоящему отличающиеся по размеру. В 2010 году Грегори С. Пол оценил длину Ornithomimus edmontonicus в 3,8 метра, а массу — в 170 кг. Один из образцов этого вида, CMN 12228, содержит бедренную кость в 46,8 см длиной. Ornithomimus velox , типовой вид рода орнитомим, основан на ископаемом материале животного меньшего размера. В то время, как голотип O. edmontonicus , CMN 8632, сохраняет длину второй пястной кости в 84 мм, тот же элемент скелета у O. velox имеет длину в 53 мм.

В палеонтологии на долгое время закрепилось убеждение, что орнитомимы, как и многие другие динозавры, были чешуйчатыми. Однако, начиная с 1995 года, было найдено несколько экземпляров орнитомимов с остатками перьевого покрова.
В 1995, 2008 и 2009 годах было обнаружено три образца Ornithomimus edmontonicus с признаками перьев. Две взрослых особи с углеродистыми следами на нижних частях передних конечностей, указывающими на прежнее присутствие перьевых очинов , которые были до 5 см длиной. Также были обнаружены остатки волосяных нитей, охватывающих по кругу охвостье, шею и ноги. Тот факт, что следы перьев обнаружились в песчанике , где, как прежде считалось, они не могли сохраняться, поднял вопрос о возможности нахождения подобных структур с более тщательной подготовкой будущих образцов. Исследователи, изучавшие образцы в 2012 году, пришли к выводу, что тела орнитомимов были покрыты пышными перьями на всех стадиях роста животных, но только взрослые особи имели оперённые крылоподобные структуры. Это натолкнуло учёных на мысль, что данные структуры служили орнитомимам для брачных игр . В 2014 году Кристиан Фот и другие утверждали, что доказательства были недостаточными, чтобы сделать вывод о том, что оперение передних конечностей орнитомимов было обязательно перьями. Учёный сослался на то, что одноволоконные перья крыла казуаров , скорее всего, оставят подобные следы .

Четвёртый оперённый образец, на этот раз из нижней части формации , описали в октябре 2015 года Аарон ван дер Реест, Алекс Вулф и Фил Карри. Это первый экземпляр орнитомима, сохранивший перья вдоль хвоста. Перья, пусть разбитые и перекошенные, имели сильное сходство в структуре и по расположению на теле с теми, которые есть у современных страусов. Отпечатки кожи у образца 2015 года также сохранились и показывали голую кожу от середины бедра до ступней, а также складку кожи, которая соединяла бедро и торс. Эта последняя структура аналогична той, которая имеется у современных птиц, в том числе страусов, но была расположена у орнитомима прямо над коленом, в отличие от птиц .
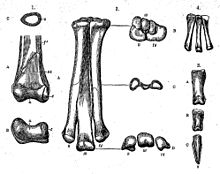

История классификации орнитомима и классификации орнитомимид в целом весьма сложна. Типовой вид, Ornithomimus velox , впервые назвал в 1890 году Отниел Чарлз Марш на основе синтипов YPM 542 и YPM 548, частичных передней и задней конечностей, обнаруженных 30 июня 1889 года Джорджем Лаймоном Кэнноном в Денверской формации в Колорадо . Родовое наименование означает «птичий имитатор», или «похожий на птицу». Видовой эпитет переводится с латинского как «быстрый» . Одновременно Марш назвал два других вида: Ornithomimus tenuis , на основе образца USNM 5814, и Ornithomimus grandis . Оба состоят из фрагментарных окаменелостей, найденных Джоном Беллом Хэтчером в штате Монтана . Эти экземпляры ныне расцениваются как материал, относящийся к тираннозавридам . Сначала Марш причислил орнитомима к подотряду орнитоподов , но изменил своё мнение после обнаружения Хэтчером в Вайоминге образца USNM 4736, частичного скелета орнитомимида, который Марш назвал Ornithomimus sedens в 1892 году. По этой причине был также создан вид Ornithomimus minutus на основе образца YPM 1049, плюсневой кости , который теперь причисляют к альваресзавридам .
Шестой вид, Ornithomimus altus , был назван в 1902 году Лоуренсом Ламбом на основании образца CMN 930, задних конечностей, найденных в 1901 году в канадской провинции Альберта , но в 1916 году Генри Осборн выделил его в самостоятельный род Struthiomimus . В 1920 году Чарльз Гилмор дал название Ornithomimus affinis для вида Dryosaurus grandis (Lull, 1911) на основе неопределённого материала. В 1930 году Лорис Расселл переименовал Struthiomimus brevetertius (Parks, 1926) и Struthiomimus samueli (Parks 1928) в Ornithomimus brevitertius и Ornithomimus samueli . В том же году Оливер Перри Хэй переименовал Aublysodon mirandus (Leidy, 1868) в Ornithomimus mirandus , который сегодня рассматривается как nomen dubium . В 1933 году создал вид Ornithomimus elegans , который сегодня рассматривается как принадлежащий либо роду Chirostenotes , либо роду Elmisaurus . В том же году Гилмор дал название Ornithomimus asiaticus ископаемому материалу, найденному в Монголии .
В том же 1933 году Чарльз Штернберг назвал вид Ornithomimus edmontonicus на основе почти полного скелета из формации Хорсшу, Альберта (образец CMN 8632) .
Сначала было принято называть каждого открытого орнитомимида как разновидность орнитомима. В 1960-х годах эта тенденция была ещё сильна, на что указывает факт переименования Оскаром Куном вида Megalosaurus lonzeensis (Dollo, 1903) из Бельгии в Ornithomimus lonzeensis (сегодня классифицируется как когти абелизавроида ) , а Дэйл Расселл в 1967 году переименовал Struthiomimus currellii (Parks, 1933) и Struthiomimus ingens (Parks, 1933) в Ornithomimus currellii и Ornithomimus ingens . В то же время научные работники обычно ссылались на весь материал, относящийся к орнитомимидам, как на струтиомима . Для того, чтобы разрешить эту путаницу, учёные сделали научное разделение между орнитомимом и струтиомимом; в 1972 году Дейл Расселл опубликовал морфометрическое исследование, которое показывает, какие статистические различия в некоторых пропорциях можно использовать для различия этих двух родов. Расселл пришёл к выводу, что струтиомим и орнитомим были валидными родами. У последнего Расселл признал два вида: Ornithomimus velox и Ornithomimus edmontonicus , даже если он не мог достоверно отличить его от O. velox. Вид Struthiomimus currellii он считал младшим синонимом Ornithomimus edmontonicus . Тем не менее, Расселл также интерпретировал эти данные как указывающие на то, что многие образцы не могут быть отнесены ни к струтиомиму, ни к орнитомиму. Таким образом, он создал два новых рода. Первым был Archaeornithomimus , к которому были причислены Ornithomimus asiaticus и Ornithomimus affinis , ставшие Archaeornithomimus asiaticus и Archaeornithomimus affinis соответственно. Вторым родом стал Dromiceiomimus , означающий «подобный эму ». Расселл отнёс несколько бывших видов орнитомима, названных в XX веке, включая O. brevitertius и O. ingens , к новому виду Dromiceimimus brevitertius . Он также переименовал вид Ornithomimus samueli во второй вид Dromiceiomimus : Dromiceiomimus samueli .

В 1865 году Джозеф Лейди причислил две большеберцовые кости из формации Нэйвсинк к виду Coelosaurus antiquus . Эти кости были впервые отнесены к орнитомиму в 1979 году Дональдом Бэрдом и Джоном Р. Хорнером под именем Ornithomimus antiquus . Обычно это сделало бы орнитомима младшим синонимом Coelosaurus , но Бэрд и Хорнер обнаружили, что название Coelosaurus было занято сомнительным таксоном на основе одного позвонка, названного так неизвестным автором. Сейчас известно, что это сделал Ричард Оуэн в 1854 году . Бэрд упоминает несколько других образцов из Нью-Джерси и Мэриленда, причисленных к Ornithomimus antiquus . Начиная с 1997 года, Роберт М. Салливан считал О. velox и О. edmontonicus младшими синонимами О. antiquus . Как и Рассел, он считал прежние два вида неотличимыми друг от друга, и отметил, что они оба разделяют отличительные черты с О. antiquus . Тем не менее, Дэвид Вейзхемпел в 2004 году рассматривал Coelosaurus antiquus как занимающего неопределённое положение среди прочих орнитомимозавров и, следовательно, nomen dubium .
В 1988 году Грегори С. Пол переименован Gallimimus bullatus в Ornithomimus bullatus . Подобная классификация не нашла поддержки у научного сообщества и на данный момент не используется самим Полом.

Даже после исследований Расселла различные исследователи продолжали относить все или некоторые из этих видов обратно к орнитомиму в различных комбинациях. В 2004 году Питер Маковицки, Йошицугу Кобаяши и Фил Карри изучили статистику пропорций Расселла 1972 года, чтобы повторно анализировать родственные отношения орнитомимозавров в свете обнаружения новых образцов. Они пришли к выводу, что не было никаких оснований для отделения дромицеомима от орнитомима, понижая дромицеомима до синонима О. edmontonicus . Тем не менее, они не включили в свой анализ типовой вид, Ornithomimus velox . Та же самая команда позже поддержала синонимию между дромицеомимом и О. edmontonicus во время лекции на ежегодном собрании Общества позвоночной палеонтологии , и их мнению последовало большинство поздних авторов . Команда Маковицки также считает Dromiceiomimus samueli младшим синонимом О. edmontonicus , хотя Лонгрич позже предположил, что он может принадлежать к различным неназванным видам из формации Дайносор парк, которые до сих пор не описаны .
Помимо O. edmontonicus , чьи ископаемые остатки датируются ранней маастрихской эпохой , два других возможно валидных вида датируются этим же периодом. О. sedens был назван Маршем в 1891 году на основе частичных ископаемых остатков, найденных в формации Ланс в Вайоминге , год спустя после описания O. velox . В своём пересмотре орнитомимидов 1972 года Дейл Расселл не указал, к какому роду принадлежит этот таксон, но предположил, что он мог занимать промежуточное положение между струтиомимом и дромицеомимом. В 1985 году он считал его разновидностью орнитомима . Несмотря на то, что с тех пор вид относят к Struthiomimus sedens , на основе полных образцов из Монтаны (а также некоторых фрагментов из Альберты и Саскачевана ), они все же должны быть описаны и сравнены с голотипом O. sedens .
Другой вид — оригинальный типовой вид O. velox , поначалу известный по весьма ограниченным ископаемым остаткам. Дополнительные образцы, отнесённые к этому виду, были обнаружены в Денверской формации и формации Феррис в Вайоминге . Один образец O. velox (MNA P1 1762A) из формации Каипаровитс, Юта , был описан в 1985 году . Повторная оценка образца, проведённая Линдси Занно и её коллегами в 2010 году, ставит под сомнение его причисление к O. velox , и возможно, к роду орнитомим в целом . Это заключение было подтверждено в 2015 году повторным описанием O. velox , которое показало, что только голотип образца можно с уверенностью отнести к этому виду. Авторы этой работы предварительно сослались на образец из формации Каипаровитс как на Ornithomimus sp. , наряду со всеми экземплярами из формации Дайносор парк .
В 1890 году Марш отнёс орнитомима к семейству орнитомимид , и классификация с тех пор не изменилась. Современные кладистические исследования показывают отдельную позицию среди орнитомимид; тем не менее, туда был включён только O. edmontonicus . Отношения между О. edmontonicus, О. velox и О. sedens не были опубликованы.
Приведённая ниже кладограмма основана на исследовании Сюй Сина и его коллег 2011 года .
| Ornithomimidae |
|
||||||||||||||||||||||||||||||||||||

Способ питания орнитомима всё ещё обсуждается. Являясь тероподами, орнитомимиды могли быть плотоядными, но их форма тела подходит и для частично или в значительной степени растительноядного образа жизни. Подходящая пища могла включать насекомых, ракообразных, плоды, листья, ветви, яйца и мясо ящериц и мелких млекопитающих .
Орнитомим имел ноги, которые кажутся пригодными для быстрого передвижения, с большеберцовой костью на 20 % длиннее, чем бедро. Большие глазницы предполагают острое зрение, а также позволяют предположить, что орнитомимы вели преимущественно ночной образ жизни .


