Ламинин
- 1 year ago
- 0
- 0

Нетрины — белки, участвующие в аксональном наведении . Название взято от санскритского слова «нетр» — «тот, кто направляет». Отмечается высокая консервативность генов нетринов среди животных — от нематод и плодовых мушек до лягушек и мышей. Структурно нетрины напоминают ламинины . Помимо регулирования нейромиграции в растущем мозге , нетрины играют роль и за его пределами, регулируя адгезию, подвижность, пролиферацию, дифференциацию и выживание клеток.
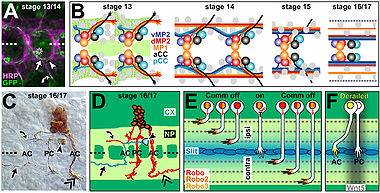
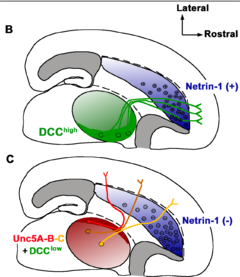
От концентрации нетринов зависит направление роста аксонов. Хотя механизм управления аксонами понят не полностью, известно, что сигнал к росту в заданном направлении опосредован UNC-40/DCC рецепторами поверхности клеток, а сигнал, означающий запрет роста в данном направлении - UNC 5 рецепторами. Также нетрины выступают в роли факторов роста , стимулируя рост клеток-мишеней. У мышей, лишённых нетринов, не может сформироваться гиппокампальная спайка или мозолистое тело .
Предлагаемая модель активности нетринов в развивающемся человеческом эмбрионе состоит в том, что нетрины производятся вентральной пластинкой нервной трубки и воспринимаются белками, находящимися на аксонов , принадлежащих нейронам развивающегося организма. Тела этих нейронов остаются на своих местах, в то время как их аксоны растут в направлении, указанном нетринами, в конечном счёте соединяясь с другими нейронами эмбрионального мозга и образуя с ними синапсы . Некоторые исследования говорят о том, что новые аксоны часто следуют уже проложенными путями (за более ранними аксонами), а не управляются нетринами.
Все нетрины состоят примерно из 600 аминокислот и имеют массу около 70 000 а. е. м.
Первыми в 1990 году были описаны нетрины нематоды Caenorhabditis elegans , названные UNC 6. Первые гомологичные белки позвоночных были открыты в 1994 году. Они руководили аксонами спинного мозга грызунов. В 2009 году было описано ещё 5 нетринов.
Нетрины в основном представлены секретируемыми белками, служащими двухфункциональными сигналами: привлечение одних нейронов и отталкивание других во время развития нервной ткани. Они обильны на срединной линии тела у всех животных, обладающих двусторонней симметрией, и могут служить сигналами, посылаемыми на большое и малое расстояние во время образования нервной ткани. В зависимости от того, хотят они привлечь аксоны или отпугнуть, нетрины взаимодействуют с DCC или UNC 5 рецепторами. Вторичная структура нетринов очень стабильна. Несколько областей нетрина в N-концевой части гомологичны ламимину. В участке, прилегающем к С-концу, наблюдается наиболее разнообразное строение у разных нетринов. С-конец содержит различные аминокислоты, позволяющие нетрину взаимодействовать с разными белками во внеклеточном матриксе или на поверхности клеток. Структурные и функциональные различия концевых участков позволили идентифицировать различные виды нетринов, такие, как нетрин-1, нетрин-3 и нетрин-G.
Нетрин-1 найден в нейроэпителиальных клетках вентральной пластинки и в клетках вентральной области спинного мозга, а также в других областях организма, включая соматическую мезодерму, поджелудочную железу и сердечную мышцу. Его главные функции - управление аксонами, передвижением нейронов и морфогенезом ветвящихся структур. Мыши с мутациями в гене нетрина-1 испытывали нехватку в клетках переднего мозга и комиссуральных аксонах нейронов спинного мозга. Нетрин-3 отличается от других нетринов. Его много в периферической нервной системе в сенсорных, моторных и вставочных нейронах и мало в центральной нервной системе. Исследования нетрина-3 показали пониженную способность взаимодействия с DCC рецепторами по сравнению с нетрином-1, из чего можно заключить, что он работает через другие рецепторы. Нетрин-G - секретируемый, но он остаётся связанным с внеклеточной поверхностью через гликозилфосфатидилинозитол (ГФИ-якорь). Нетрин-G преобладает в центральной нервной системе в таких участках, как таламус и митральные клетки обонятельной луковицы. Он взаимодействует не с DCC и UNC-5, а с лигандом NGL-1. Два вида нетрина-G, нетрин-G1 и нетрин-G2, обнаружены только у позвоночных. Считается, что их появление облегчило строительство сложного головного мозга позвоночных.
Конусы роста, расположенные на конце аксонов, отвечают за удлинение аксонов во время развития нейронов при эмбриогенезе. Удлинение происходит в ответ на трофические и иные факторы, присутствующие в окружающей среде. Нетрины - одни из таких факторов. Исследования многих организмов (включая мышей, крыс, цыплят, нематод Caenorhabditis elegans и дрозофил Drosophila melanogaster) показали, что нетрины бифункциональны: они могут выступать и в роли аттрактантов, и в роли репеллентов в отношении аксонального роста, а также могут действовать как на ближние клетки, так и на дальние.
Исследования центральной нервной системы цыплят и грызунов показали, что нетрин-1 является чрезвычайно важной частью системы управления ростом аксонов. Наиболее важно то, что было замечено секретирование нетрина-1 специализированными клетками эмбрионального головного мозга. В брюшной средней линии нетрин-1 наиболее ярко выражен, и его концентриция уменьшается по мере движения в дорсальном направлении. Дополнительные исследования мышей, испытывающих недостаток нетрина, показали, что когда нетрин связывается с DCC рецептором на конусе роста аксона, стимулируется реакция привлечения аксона. Это также подтверждается отсутствием у наблюдаемых мышей развития вентральной спаянности спинного мозга. Подобные результаты наблюдались в экспериментах с гомологом нетрина-1, обнаруженным у нематод Caenorhabditis elegans. Тот же самый градиент концентрации белка наблюдается в эпидермальных клетках развивающегося червя. Данные свидетельствуют, что этот градиент важен для дальнего действия UNC-6 в управлении начальным передвижением аксонов к брюшной средней линии,и что привлекающая реакция опосредована UNC-40. Когда аксоны достигают средней линии, концентрация UNC-6 резко уменьшается, сигнализируя о том, что после общего управления ростом в спинно-брюшном направлении UNC-6 далее вовлечён в направление аксонов к более специальным местам. Недавно ученые описали много клеточных механизмов, с помощью которых нетрин-1, связанный с DCC, привлекает аксоны через как минимум три сигнальных пути. На начальном этапе всех трёх путей нетрин-1 вызывает гомодимеризацию DCC, который запускает сигнальный каскад, приводящий к хемоатрактации.
Исследования DCC позвоночных и UNC-40 Caenorhabditis elegans показали, что отталкивающая реакция аксонов всегда связана с UNC5 рецепторами. В том же самом градиенте брюшной средней линии, обсуждённом выше, нетрин-1 действует как хеморепеллент для аксонов моторных нейронов, направляя их рост дорзально (дальше от брюшной средней линии). Большое количество дефектов наблюдалось у Caenorhabditis elegans с мутировавшими UNC-40; однако ошибки в перемещениях аксонов были более сильно выражены у нематод с мутациями в UNC-5 генах, указывая на связь гомолога нетрина-1 UNC-6 с UNC-5 рецепторами, которые одни могут отпугивать аксоны. Отталкивание в хемоотражающих системах малой дальности действия и позвоночных, и беспозвоночных происходит через UNC-5 рецепторы, в то время как для систем дальнего действия требуются и DCC (UNC-40 у Caenorhabditis elegans) и UNC-40.