.
.
 .
.
 ,
,
 .
.
 .
.
 .
.
 .
.
 .
.
![{\displaystyle {\ce {2 NO ->[BaO, >700^oC] N2 + O2}}}](/images/000/035/35792/19.svg?rand=489666) .
.

Оксид празеодима(II)
- 1 year ago
- 0
- 0

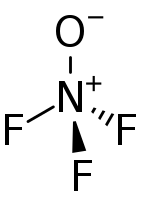
Окси́д азо́та(II) ( мон(о)оксид азота , окись азота , нитрозил-радикал ) NO — несолеобразующий оксид азота . Молекула года (1992)
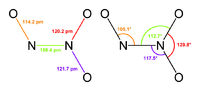
Наличие неспаренного электрона обусловливает склонность NO к образованию слабосвязанных димеров N 2 O 2 . Это непрочные соединения с Δ H ° димеризации около 17 кДж/моль. Жидкий оксид азота(II) на 25 % состоит из молекул N 2 O 2 , а твёрдый целиком состоит из них.
Оксид азота(II) — единственный из оксидов азота, который можно получить непосредственно из свободных элементов соединением азота с кислородом при высоких температурах (1200—1300 °C) или в электрическом разряде. В природе он образуется в атмосфере при грозовых разрядах (тепловой эффект реакции −180,9 кДж):
и тотчас же реагирует с кислородом :
При понижении температуры оксид азота(II) разлагается на азот и кислород, но если температура падает резко, то не успевший разложиться оксид существует достаточно долго: при низкой температуре скорость распада невелика. Такое резкое охлаждение называется «закалкой» и используется при одном из способов получения азотной кислоты .
В лаборатории его обычно получают взаимодействием 31 % HNO 3 с некоторыми металлами , например, с медью :
Более чистый, не загрязнённый примесями NO можно получить по реакциям
Промышленный способ основан на окислении аммиака при высокой температуре и давлении при участии Pt , Rh , Cr 2 O 3 (как катализаторов ):
Получение NO является одной из стадий получения азотной кислоты .
В нормальных условиях NO представляет собой бесцветный газ. Плохо растворим в воде. Имеет плотность 1,3402 кг/м³ . Сжижается с трудом; в жидком и твёрдом виде имеет голубой цвет.
При комнатной температуре и атмосферном давлении происходит окисление NO кислородом воздуха:
В результате смесь газов приобретает коричневый цвет.
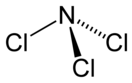
Для NO характерны также реакции присоединения галогенов с образованием нитрозилгалогенидов, в этой реакции NO проявляет свойства восстановителя с образованием нитрозилхлорида :
В присутствии более сильных восстановителей NO проявляет окислительные свойства:
При температуре свыше +700 °C в присутствии оксида бария разлагается:
.
С водой не реагирует, является несолеобразующим оксидом.
Оксид азота (II) — ядовитый газ с удушающим действием.

Оксид азота является одним из немногих известных газотрансмиттеров и, кроме того, является также химически высокореактивным свободным радикалом, способным выступать как в роли окислителя, так и в роли восстановителя. Окись азота является ключевым вторичным посредником в организмах позвоночных и играет важную роль в межклеточной и внутриклеточной передаче сигнала и, как следствие, во множестве биологических процессов. Известно, что окись азота производится практически всеми типами живых организмов, от бактерий, грибов и растений до клеток животных.
Окись азота, первоначально известная под именем эндотелиального сосудорасширяющего фактора (химическая природа которого тогда ещё была не известна) синтезируется в организме из аргинина при участии кислорода и НАДФ ферментом синтазой оксида азота . Восстановление неорганических нитратов также может быть использовано для производства организмом эндогенной окиси азота. Эндотелий кровеносных сосудов использует окись азота в качестве сигнала окружающим гладкомышечным клеткам расслабиться, что приводит к вазодилатации и увеличению кровотока. Окись азота является высокореактивным свободным радикалом со временем жизни порядка нескольких секунд, но при этом обладает высокой способностью к проникновению сквозь биологические мембраны. Это делает окись азота идеальной сигнальной молекулой для кратковременного аутокринного (внутри клетки) или паракринного (между близко расположенными или соседними клетками) обмена сигналами.
Независимо от активности синтазы оксида азота, существует и другой путь биосинтеза окиси азота, так называемый нитрат-нитрит-оксидный путь, состоящий в последовательном восстановлении пищевых нитратов и нитритов, получаемых из растительной пищи. Было показано, что богатые нитратами овощи, в особенности листовая зелень , такая, как шпинат и руккола , а также свёкла , способны повышать уровень эндогенной окиси азота и обеспечивать защиту миокарда от ишемии, а также снижать артериальное давление у лиц с предрасположенностью к артериальной гипертензии или начинающимся развитием АГ. Для того, чтобы организм мог производить окись азота из нитратов пищи по нитрат-нитрит-оксидному пути, сначала обязательно должно произойти восстановление нитратов до нитритов с помощью сапрофитных бактерий (бактерий-комменсалов), которые обитают во рту. Мониторинг содержания окиси азота в слюне позволяет обнаружить биотрансформацию растительных нитратов в нитриты и окись азота. Повышение уровня окиси азота в слюне наблюдается при диетах, богатых листовой зеленью. В свою очередь, листовая зелень — часто важнейший компонент многих антигипертензивных и «сердечных» диет, разработанных для лечения гипертонической болезни, ишемической болезни сердца, сердечной недостаточности.
Выработка окиси азота повышена у людей, живущих в горах, особенно на больших высотах. Это способствует приспособлению организма к условиям пониженного парциального давления кислорода и уменьшению вероятности гипоксии за счёт увеличения кровотока как в лёгких, так и в периферических тканях. Известные эффекты окиси азота включают в себя не только вазодилатацию, но и участие в нейротрансмиссии в качестве газотрансмиттера, и активацию роста волос, и образование реактивных промежуточных продуктов обмена, и участие в процессе эрекции пениса (благодаря способности окиси азота расширять сосуды полового члена). Фармакологически активные нитраты, такие, как нитроглицерин , амилнитрит , , реализуют своё вазодилатирующее, антиангинальное (антиишемическое), гипотензивное и спазмолитическое действие благодаря тому, что из них в организме образуется окись азота. Вазодилатирующее гипотензивное лекарство миноксидил содержит остаток NO и может работать, кроме всего прочего, ещё и как агонист NO. Аналогично, силденафил и подобные ему препараты способствуют улучшению эрекции преимущественно за счёт того, что усиливают работу связанного с NO сигнального каскада в половом члене.
Окись азота способствует поддержанию гомеостаза сосудов, вызывая расслабление гладких мышц стенок сосудов и угнетая их рост и утолщение интимы сосудов (гипертензивное ремоделирование сосудов), а также угнетая адгезию и агрегацию тромбоцитов и адгезию лейкоцитов к эндотелию сосудов. У больных с атеросклерозом сосудов, сахарным диабетом или гипертензией часто имеются признаки нарушения обмена оксида азота или нарушения во внутриклеточных каскадах передачи сигнала от оксида азота.
Было также показано, что высокое потребление соли снижает образование окиси азота у больных с гипертонической болезнью, хотя биодоступность окиси азота не меняется, остаётся прежней.
Окись азота также образуется в процессе фагоцитоза такими способными к фагоцитозу клетками, как моноциты , макрофаги , нейтрофилы , как часть иммунного ответа на вторжение чужеродных микроорганизмов (бактерий, грибков и др.). Клетки, способные к фагоцитозу, содержат индуцируемую синтазу оксида азота (iNOS), которая активируется γ-интерфероном или сочетанием фактора некроза опухоли со . С другой стороны, β-трансформирующий фактор роста (TGF-β) оказывает сильное угнетающее действие на активность iNOS и биосинтез оксида азота фагоцитами. Интерлейкины 4 и 10 оказывают слабое угнетающее действие на активность iNOS и биосинтез оксида азота соответствующими клетками. Таким образом, иммунная система организма обладает способностью регулировать активность iNOS и доступный фагоцитам арсенал средств иммунного ответа, что играет роль в регуляции процессов воспаления и силы иммунных реакций. Оксид азота секретируется фагоцитами в процессе иммунного ответа в качестве одного из свободных радикалов и является высокотоксичным для бактерий и внутриклеточных паразитов, включая лейшманий и малярийных плазмодиев. Механизм бактерицидного, противогрибкового и антипротозойного действия оксида азота включает в себя повреждение ДНК бактерий, грибков и простейших и повреждение железосодержащих белков с разрушением комплексов железа с серой и образованием нитрозилов железа.
В ответ на это многие патогенные бактерии, грибки и простейшие эволюционно развили механизмы устойчивости к образующемуся в процессе фагоцитоза оксиду азота или механизмы его быстрого обезвреживания. Поскольку повышение образования эндогенного оксида азота является одним из маркеров воспаления и поскольку эндогенный оксид азота может оказывать провоспалительное действие при таких состояниях, как бронхиальная астма и бронхообструктивные заболевания, в практической медицине наблюдается повышенный интерес к возможному использованию анализа на содержание оксида азота в выдыхаемом воздухе в качестве простого дыхательного теста при заболеваниях дыхательных путей, сопровождающихся их воспалением. Пониженные уровни эндогенного оксида азота в выдыхаемом воздухе были обнаружены у курильщиков и у велосипедистов, подвергающихся воздействию загрязнения воздуха. В то же время в других популяциях (то есть не среди велосипедистов) с воздействием загрязнения воздуха ассоциировалось повышение уровня эндогенного оксида азота в выдыхаемом воздухе.
Эндогенный оксид азота может привносить свой вклад в повреждение тканей при ишемии и последующей реперфузии, поскольку в процессе реперфузии может образовываться избыточное количество оксида азота, который может реагировать с супероксидом или пероксидом водорода и образовывать сильный и токсичный окислитель, повреждающий ткани — пероксинитрит . Напротив, при отравлении паракватом вдыхание оксида азота способствует повышению выживаемости и лучшему восстановлению больных, поскольку паракват вызывает образование в лёгких больших количеств супероксида и пероксида водорода, снижение биодоступности NO вследствие его связывания с супероксидом и образования пероксинитрита и угнетение активности синтазы оксида азота.
У растений эндогенный оксид азота может производиться одним из четырёх способов:
У растений эндогенный оксид азота также является сигнальной молекулой (газотрансмиттером), способствует снижению или предотвращению оксидативного стресса клеток, а также играет роль в защите растений от патогенных микроорганизмов и грибков. Было показано, что воздействие низких концентраций экзогенного оксида азота на срезанные цветы и другие растения увеличивает продолжительность времени до их увядания, пожелтения и осыпания листьев и лепестков.
Два важнейших механизма, при помощи которых эндогенный оксид азота проявляет своё биологическое действие на клетки, органы и ткани — это S-нитрозилирование тиоловых соединений (включая тиоловые группы серосодержащих аминокислот , таких, как цистеин ) и нитрозилирование ионов переходных металлов. S-нитрозилирование означает обратимое преобразование тиоловых групп (например, цистеиновых остатков в составе молекул белков) в S-нитрозотиолы (RSNO). S-нитрозилирование является важным механизмом динамической, обратимой посттрансляционной модификации и регуляции функций многих, если не всех, основных классов белков. Нитрозилирование ионов переходных металлов подразумевает связывание NO с ионом переходного металла, такого, как железо , медь , цинк , хром , кобальт , марганец , в том числе с ионами переходных металлов в составе простетических групп или активных каталитических центров металлоферментов. В этой роли NO является нитрозильным лигандом . Типичные случаи нитрозилирования ионов переходных металлов включают в себя нитрозилирование гем -содержащих белков, таких, как цитохром , гемоглобин , миоглобин , что приводит к нарушению функции белка (в частности, невозможности гемоглобина выполнять свою транспортную функцию, или инактивации фермента). Особенно важную роль играет нитрозилирование двухвалентного железа, поскольку связывание нитрозильного лиганда с ионом двухвалентного железа особенно сильное и приводит к образованию очень прочной связи. Гемоглобин является важным примером белка, функция которого может изменяться под влиянием NO обоими способами: NO может как непосредственно связываться с железом в составе гема в реакции нитрозилирования, так и образовывать S-нитрозотиолы при S-нитрозилировании серосодержащих аминокислот в составе гемоглобина.
Таким образом, существует несколько механизмов, при помощи которых эндогенный оксид азота оказывает двоякое влияние на биологические процессы в живых организмах, клетках и тканях. Эти механизмы включают окислительное нитрозилирование железосодержащих и других металлосодержащих белков, таких, как рибонуклеотид-редуктаза, аконитаза, активацию растворимой гуанилатциклазы с повышением образования цГМФ , стимуляцию АДФ-зависимого рибозилирования белков, S-нитрозилирование сульфгидрильных (тиоловых) групп белков, приводящее к их посттрансляционной модификации (активации либо инактивации), активацию регулируемых факторов транспорта железа, меди и других переходных металлов. Было также показано, что эндогенный оксид азота способен активировать ядерный фактор транскрипции каппа (NF-κB) в мононуклеарных клетках периферической крови. А известно, что NF-κB является важным фактором транскрипции в регуляции процессов апоптоза и воспаления, и в частности важным фактором транскрипции в процессе индукции экспрессии гена индуцируемой синтазы оксида азота. Таким образом, продукция эндогенного оксида азота саморегулируется — повышение уровня NO угнетает дальнейшую экспрессию индуцируемой синтазы оксида азота и предотвращает чрезмерное повышение её уровня и чрезмерное повреждение тканей организма хозяина в процессе воспаления и иммунного ответа.
Известно также, что вазодилатирующее действие оксида азота опосредуется в основном через стимуляцию им активности растворимой гуанилатциклазы, являющейся гетеродимерным ферментом, активирующимся при нитрозилировании. Стимуляция активности гуанилатциклазы приводит к накоплению циклического ГМФ. Увеличение концентрации в клетке циклического ГМФ приводит к повышению активности протеинкиназы G. Протеинкиназа G, в свою очередь, фосфорилирует ряд важных внутриклеточных белков, что приводит к обратному захвату ионов кальция из цитоплазмы во внутриклеточные хранилища и к открытию активируемых кальцием калиевых каналов . Снижение концентрации ионов кальция в цитоплазме клетки приводит к тому, что киназа лёгкой цепи миозина, активируемая кальцием, теряет активность и не может фосфорилировать миозин, что приводит к нарушению образования в молекуле миозина «мостиков» и нарушению его свёртывания в более компактную структуру (сокращения), а следовательно и к расслаблению гладкомышечной клетки. А расслабление гладкомышечных клеток стенок сосудов ведёт к расширению сосудов (вазодилатации) и увеличению кровотока.