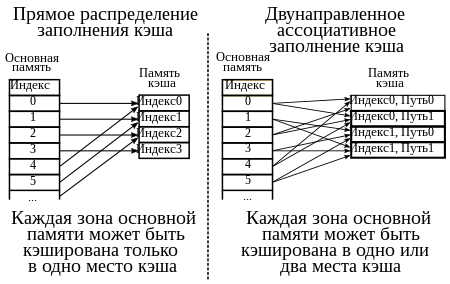
Алгоритмы кэширования
- 1 year ago
- 0
- 0

Питека́нтроп (от др.-греч. πίθηκος — обезьяна и ἄνθρωπος — человек) — ископаемая внутривидовая разновидность людей , известная по окаменелостям с острова Ява ( Индонезия ) и часто рассматриваемая как подвид Homo erectus erectus в рамках вида Человек прямоходящий .
Впервые питекантроп был описан Эженом Дюбуа в отчёте 1892 года и монографии 1894 года, в которой было введено новое родовое имя Pithecanthropus . Окаменелости, на основе которых был описан вид Pithecanthropus erectus , включали найденные у деревни Триниль черепной свод и моляр архаичного вида, а также бедренную кость более прогрессивной формы. Это противоречие вызвало широкую дискуссию среди специалистов, в итоге научное сообщество признало выводы Дюбуа . Некоторое время питекантроп считался « недостающим звеном » в эволюции человека от обезьяноподобных предков, но впоследствии, с объединением ряда региональных окаменелостей в общий вид Homo erectus , его стали рассматривать как тупиковую местную ветвь этого вида .
Для питекантропов характерен удлинённый низкий череп с гипертрофированной толщиной свода, широкой затылочной костью , покатым лбом и узким лицевым отделом с массивными челюстями. По многим признакам, таким как объём и форма черепа, форма зубов, средний рост, питекантропы занимают промежуточное положение между более ранними гомининами и человеком разумным . В рамках вида Homo erectus черты питекантропов более архаичны, чем у африканских представителей вида, находок из Китая или позднего человека прямоходящего из Индонезии, известного как явантроп .
Древность окаменелостей питекантропа неоднократно пересматривалась, оценка возраста наиболее древних костей колеблется между 1,8 и 1,3 миллиона лет назад . Время исчезновения питекантропа, произошедшего менее 900 тысяч лет назад, совпадает с периодом смены лесостепной плейстоценовой фауны на фауну дождевых лесов современного типа . Представления о материальной культуре питекантропов неполны: хотя на Яве обнаружены каменные орудия , их связь с питекантропами оспаривается. Имеются некоторые данные, свидетельствующие о том, что питекантроп использовал в качестве орудий труда раковины моллюсков и акульи зубы .
Термин «питекантроп» впервые был предложен в 1868 году немецким биологом Эрнстом Геккелем как обозначение гипотетического промежуточного звена между обезьяной и человеком. Эта промежуточная форма, по мнению Геккеля, должна была быть уже прямоходящей и иметь более развитый, чем у обезьян, мозг, однако всё ещё не владеть членораздельной речью . Геккель, один из отцов эволюционной теории , расходился во взглядах с Чарльзом Дарвином по вопросу о месте происхождения человека. Если Дарвин предполагал, что корни человечества следует искать в Африке , то Геккель придерживался мнения о более близком родстве человека с высшими обезьянами Юго-Восточной Азии .
Само открытие питекантропа связано с изысканиями нидерландского анатома Эжена Дюбуа . Этот учёный был сторонником эволюционной теории, включая идею о происхождении человека от обезьяноподобных предков. При этом он придерживался мнения, что наиболее логичным местом для поиска следов этих предков будут тропики , а конкретнее — Нидерландская Ост-Индия . Выбор этого региона для поисков был связан как с необходимостью финансирования со стороны нидерландского правительства, так и с убеждением, что высшие обезьяны Восточной Азии (в частности, гиббоны ), а также ряд ископаемых приматов этого зоогеографического региона (включая сивапитеков ) наиболее близки по своей анатомии к человеку . Последнюю идею Дюбуа, вероятно, почерпнул у своих университетских учителей, на которых значительное влияние оказали работы Геккеля .
В 1887 году Дюбуа, не добившись интереса к своим идеям в министерстве колоний, отправился в Нидерландскую Ост-Индию в статусе военного врача. В 1889 году ему удалось заручиться поддержкой колониальной администрации: он был зачислен в штат департамента просвещения, религии и промышленности (это ведомство в описываемый период возглавлял нидерландский востоковед Виллем Груневелдт , которому натуралист и подчинялся с этого момента напрямую , а 20 лет спустя назвал в его честь найденного в Индонезии ископаемого быка ). После этого Дюбуа начал поиски предка современного человека на острове Суматра , однако они оказались безрезультатными, и в 1890 году учёный перебрался на остров Ява . В ноябре того же года Дюбуа и его помощники нашли близ восточнояванской деревни Кедунг-Брубус, расположенной на нагорье Кенденг, фрагмент челюсти древнего вида, однако она была настолько неполной, что её не сочли заслуживающей дальнейшего изучения . Эта находка — правая передняя часть нижней челюсти — впоследствии стала известной как Кедунг-Брубус 1, или Питекантроп A; при этом её принадлежность данному виду была установлена лишь в 1924 году .

В августе 1891 года группа Дюбуа начала раскопки у восточнояванской деревни Триниль , расположенной в излучине реки Соло . К этому времени у учёного диагностировали малярию , он был освобождён от службы и мог распоряжаться собой, как хотел. Его первой важной находкой стал обнаруженный в сентябре окаменевший моляр . Рельеф жевательной поверхности, величина коронки и корней указывали на то, что этот зуб принадлежал или древнему человеку, или крупной человекообразной обезьяне. Дюбуа пришёл к выводу, что моляр — меньший по размерам, чем у современного человека, но с необычно длинными и широко расставленными корнями, — принадлежал обезьяне, вероятно, шимпанзе . В последующем докладе исследователь сообщал о нём как о зубе вида Anthropopithecus troglodytes (родовое имя Anthropopithecus в это время носили как шимпанзе, так и ископаемые обезьяны, кости которых нашёл Ричард Лидеккер в Индии ).
В октябре неподалёку от места находки моляра рабочие Дюбуа извлекли из земли черепную крышку с узким и покатым лбом, мощными надглазничными валиками , следами сагиттального гребня и широкой уплощённой затылочной частью. Дюбуа пришёл к выводу, что черепная крышка, как и зуб, принадлежит шимпанзе, однако отметил необычайно большие размеры черепа, соответствовавшие объёму мозга примерно в 800—850 см³ (у известных человекообразных обезьян объём мозга не превышал 610 см³ , у современного человека от 930) .

В августе 1892 года группа Дюбуа нашла у Триниля глубоко минерализовавшуюся бедренную кость со следами зубов крокодила на головке. Находка была сделана неподалёку от прежних находок и вдобавок в том же слое (по оценке Дюбуа, позже подтверждённой Л. Ж. К. ван Эсом, это были нижнечетвертичные отложения ), была окрашена в такой же шоколадно-коричневый цвет и заметно деформирована патологическим разрастанием костной ткани. По длине кости (45,5 см ) рост её обладателя определялся примерно в 170 см . Хотя первоначально Дюбуа принял кость за человеческую, при ближайшем рассмотрении становились очевидны её отличия. Сама она была более прямой, чем у современного человека или известного к тому времени неандертальца , подколенная ямка была выпуклой, а не вогнутой, у сустава нижний отдел кости расширялся резко, а не постепенно .
Форма кости, несколько отличная от привычной, всё же однозначно указывала на то, что её обладатель передвигался на двух ногах. На это же указывал и развитый узел прикрепления мускулов, характерный для прямохождения. Исходя из того, что бедренная кость принадлежала тому же существу, что и найденные ранее моляр и черепная крышка, Дюбуа сделал вывод об открытии нового примата — прямоходящего и обладающего объёмом головного мозга, достигающим 2/3 от объёма мозга современного человека . Изучая свои находки, Дюбуа применил новый тип анализа, в дальнейшем получивший название аллометрического . Сравнивая объём черепа с предполагаемым ростом существа, оцениваемым на основании длины бедренной кости, он пришёл к выводу, что их соотношение не соответствует пропорциям тела ни современных людей (в том числе страдающих от патологий, связанных с уменьшением размеров черепа), ни известных человекообразных обезьян . Существо, занимающее промежуточное положение между человеком и обезьянами, он назвал Anthropopithecus erectus — «человекообезьяна прямоходящая» .
Первоначальные выводы Дюбуа содержались в его третьем квартальном ведомственном отчёте 1892 года, содержание которого вскоре стало достоянием общественности. Продолжив сравнение найденных им костей с костями шимпанзе, гиббонов и современных людей, он в 1894 году свёл свои выводы в монографию, в которой уже напрямую назвал описываемого примата переходной формой и дал ему новое имя — Pithecanthropus erectus .
Представленные Дюбуа результаты вызвали резкую критику. Уже после публикации отчёта 1892 года его обвинили в создании химеры из костей совершенно разных особей — черепа и зуба обезьяны и бедренной кости человека. Среди критиков были издатель газеты Bataviaasch Nieuwsblad Паулус Адрианус Даум (опубликовавший насмешливую статью, подписанную «Homo erectus») и немецкий зоолог Пауль Мачи . После публикации монографии к критикам Дюбуа, предполагавшим, что он соединил кости разных биологических видов, присоединились французский антрополог Леонс-Пьер Мануврие (в то же время расценивший остатки черепной коробки как самодостаточное доказательство существования обезьяночеловека) и анатом Вильгельм Краузе . Одним из самых жёстких критиков стал Рудольф Вирхов , заявивший, что черепная крышка, найденная Дюбуа, принадлежит гиббону, а бедренная кость — современному человеку. Сомнения на этом этапе выражало большинство учёных, за исключением американского палеонтолога Отниела Марша и собственно Геккеля, чью теорию Дюбуа подтверждал своей работой .
Немедленно по возвращении в Европу Дюбуа начал защиту своих выводов, посещая научные конференции и симпозиумы, демонстрируя свои находки, публикуя дополнительную информацию о геологии и фауне Триниля. За период с 1895 по 1900 год он издал 19 публикаций, посвящённых питекантропу, а его европейские коллеги — 95. Со временем аргументы Дюбуа завоевали многочисленных сторонников, и в научном обществе сложился консенсус по поводу статуса питекантропа как «промежуточного звена». Уже в 1899 году в новом журнале Zeitschrift für Morphologie und Anthropologie его редактор, анатом Густав Швальбе , представил эволюционное древо , связывающее человекообразных обезьян, питекантропа, неандертальца и современного человека . В то же время выводы Дюбуа пытались опровергнуть и спустя несколько десятилетий. Так, в 1923 году П. А. Минаков деминерализовал человеческий череп и подверг его деформации, пытаясь таким образом объяснить необычную форму черепной коробки из Триниля. Дюбуа, однако, указал, что ни кости питекантропа, ни сотни других костей из тех же слоёв не были ни деминерализованными, ни деформированными .

В 1930-е годы были сделаны многочисленные находки в Китае, некоторое время известные как Sinanthropus pekinensis . В 1939 году Густав фон Кёнигсвальд и Франц Вейденрейх продемонстрировали морфологическое родство между известными костями питекантропа и синантропа, и они были вначале объединены в один род ( Pithecanthropus ), а в 1950-е годы переклассифицированы как Homo erectus . Последнее было сделано на основе аргументов систематика Эрнста Майра , продемонстрировавшего неоправданную разницу в подходах к таксономии ископаемых гоминид и современных животных. Майр доказывал, что различия между питекантропом и синантропом не выходят за рамки подвидовых . Яванский человек прямоходящий с тех пор часто рассматривается как номинальный подвид Homo erectus erectus .
Также в 1930-е годы в бассейне реки Соло на Яве (в том числе в Сангиране и у расположенного недалеко оттуда посёлка ) фон Кёнигсвальдом были раскопаны дополнительные кости гоминида , чертами сходного с питекантропом, но более молодого: возраст самых поздних костей составляет менее 0,1 миллиона лет (по некоторым оценкам, до 27 тысяч лет ). Первоначально этот гоминид был описан как Homo (Javanthropus) soloensis . Явантропов из Нгандонга при схожем с питекантропом архаичном строении черепа, указывающем на преемственность, отличает несколько больший объём мозга — в среднем около 1150 см³ . Соответственно, многими учёными явантропы в XXI веке, подобно синантропам, рассматриваются как представители вида Человек прямоходящий , в том числе в рамках подвида H. erectus erectus . Другие авторы продолжают рассматривать их как отдельный вид Homo soloensis либо выделяют наиболее молодые фоссилии в качестве костей раннего сапиенса — Homo sapiens soloensis . В начале XXI века был описан найденный в районе Самбунгмачан в Центральной Яве черепной свод, по ряду характеристик занимающий промежуточное положение между фоссилиями классического и нгандонгского человека прямоходящего. Эта находка указывает на возможность, что явантропы не только входят с питекантропами в один вид, но и являются их потомками .
В 1936 году на северо-востоке Явы близ города Моджокерто экспедицией фон Кёнигсвальда был найден детский череп архаичного вида . Когда фон Кёнигсвальд объявил, что найден новый череп питекантропа, с наиболее резкой критикой этого мнения выступил сам Дюбуа, даже обвинивший коллегу в некорректной реконструкции черепа из фрагментов . Дюбуа как главный редактор журнала, в котором было опубликовано сообщение фон Кёнигсвальда, самовольно изменил родовое название Pithecanthropus на Homo , и в итоге в систематику вошёл вид Homo modjokertensis , однако в дальнейшем фоссилии из Моджокерто всё же включили в вид Homo erectus . Морфологические и хронологические отличия между ранними представителями человека прямоходящего на Яве и питекантропа частью палеоантропологов рассматриваются как свидетельство их принадлежности к разным подвидам одного вида . Согласно другой точке зрения, пересечения морфологических характеристик архаичного и классического человека прямоходящего с Явы дают основания рассматривать их как представителей одной группы в составе вида Homo erectus , отличной от явантропа . На это же указывает и близость по времени отдельных окаменелостей в регионе, образующих практически непрерывающуюся хронологическую последовательность на протяжении отрезка в 700 тысяч лет . Возможно также, что данный отрезок в действительности был значительно более коротким. Это связано с тем, что в ряде публикаций XXI века оценка возраста фоссилий из Моджокерто в 1,8 миллиона лет оспаривается как завышенная , и вместо этого предлагается примерный возраст в 1,49 миллиона лет, более близкий к датировке других ранних остатков гоминин в Центральной и Восточной Азии .
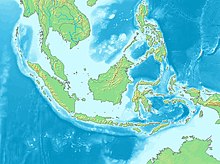
По оценкам палеоантропологов, первая волна миграции Homo erectus могла достигнуть территории современного Китая уже 2,1 миллиона лет назад, а 1,8 миллиона лет назад в Юго-Западную Азию из Африки пришла уже вторая волна. Эти данные означают, что индонезийский человек прямоходящий, чьи наиболее ранние окаменелости в зависимости от источника датируются 1,8—1,3 миллиона лет назад, мог быть как потомком второй волны миграции, пошедшей по отличному от первой маршруту, так и потомком древнейшего Homo erectus из Китая, предшествовавшего синантропу . По всей видимости, проникновение человека на субконтинент Сунда , остатками которого являются острова современной Индонезии, произошло в один из периодов глобального похолодания. Поскольку из-за ухода океанской воды в ледники уровень Мирового океана понизился, стал возможен переход из материковой Восточной Азии на территорию Сунды посуху .
Помимо существующей классификации, согласно которой питекантроп является подвидом внутри вида Homo erectus , среди части палеоантропологов, начиная с 1960-х годов, существует точка зрения, согласно которой сам этот вид невалиден. Согласно этой позиции, его подвиды должны быть включены в вид Homo sapiens , плавно эволюционировавший на протяжении плейстоцена . В этом случае таксономически питекантроп наряду с синантропом может рассматриваться как представитель подвида Homo sapiens erectus . Эта теория восходит к идеям полицентризма в человеческой эволюции, выдвинутым ещё в 1940-е годы Вейденрейхом; этот автор полагал, что коренное население Австралии , как минимум, частично происходит от архаичных людей Индонезии (питекантропа и явантропа) .
Хотя на протяжении некоторого времени после открытия питекантроп рассматривался как «недостающее звено» в эволюции неандертальца и современного человека от обезьяноподобных предков, позже в антропологии утвердилось мнение, что прямым предком человека разумного индонезийский человек прямоходящий, по-видимому, не был . Согласно господствующей на начало XXI века точке зрения, основной процесс эволюции человека проходил в Африке. Даже если человек прямоходящий из Китая и принимал какое-то участие в эволюции человека разумного (как предполагают сторонники теории полицентризма), островная восточноазиатская популяция определённо была тупиковой ветвью развития, которую в дальнейшем вытеснили более развитые представители рода Homo .

Голотипом для вида Homo erectus в целом является крыша черепа , описанная Дюбуа в 1894 году и известная как Trinil 2. Этот образец предоставил первичную основу для выделения ключевых морфологических характеристик, таких как объём черепной коробки, низкий свод и неразрывность надглазничного валика . В то же время в силу своей неполноты голотип не позволяет определить многие важные характеристики строения черепа, которые впоследствии были установлены на основании дополнительных находок . Общее количество таких находок достаточно велико: только в центральнояванском Сангиране обнаружено более 80 окаменелостей, принадлежащих Homo erectus и датируемых периодом с 1,6 до 0,9 миллиона лет назад . К числу наиболее важных находок из Сангирана относится так называемый Pithecanthropus VIII (или Sangiran 17 ) — череп с хорошо сохранившейся коробкой (в которой не хватает только костей левой скулы), сохранившимися лицевыми костями, включая ряд зубов, и достаточно полным основанием . Sangiran 17, обнаруженный в 1969 году, остаётся самым сохранным черепом человека прямоходящего из найденных в Восточной Азии .
Оригинальное описание вида в работах Дюбуа во многом опиралось также на другую находку нидерландского учёного — бедренную кость Trinil 1. В период полемики вокруг яванских окаменелостей эта кость играла ключевую роль в доказательстве того, что её обладатель был прямоходящим . Однако более поздние находки бедренных костей, отождествляемых с питекантропом, в том числе и при раскопках в самом Триниле, существенно отличались от самой первой. Эти различия отмечаются также при сравнении Trinil 1 с бедренными костями архаичных представителей рода Homo из Африки. У неё, в частности, отсутствуют такие ключевые характеристики, как относительно длинная шейка и низкое положение минимальной срединно-боковой ширины тела кости. Таким образом, существует высокая вероятность, что эта бедренная кость в действительности принадлежит значительно более позднему виду и что её первоначальная датировка и включение в описание Pithecanthropus erectus ошибочны .
Питекантроп обладал невысоким ростом, прямохождением и архаичным строением черепа . Средний рост яванских представителей Homo erectus оценивается в 160 см — больше, чем у их родичей из Китая и Дманиси , но меньше, чем у африканских представителей этого вида, для которых оценки колеблются от 160 до 180 см . Человек прямоходящий как вид в основном определяется по характеристикам черепа и зубного аппарата. Для этих ранних людей характерен длинный и относительно низкий, угловатый череп со скошенным лбом, с объёмом мозга от 700 до 1200 см³ , возрастающим по мере приближения к современности . Объём мозга у классических представителей вида с Явы находится в средней части этого диапазона и варьирует от 800 см³ для черепа Sangiran 2 (одного из древнейших обнаруженных на Яве ) до чуть более чем 1000 для черепов Sangiran 10 и Sangiran 17 .
Череп расширяется книзу, часто достигая максимальной ширины в районе надсосцевидного гребня (выступа чешуйчатой части височной кости , соединяющего верхний край скулового отростка с нижней височной линией). Кости свода черепа в целом толстые. Некоторые кости черепа гипертрофированы, в их числе выделяются ясно выраженный надглазничный валик, обычно непрерывный и утолщающийся по краям, височные и угловые (на теменной кости ) валики и центрально-осевые ( сагиттальный и лобный) гребни. Основание черепа менее выпуклое, чем у человека разумного, но не такое плоское, как у крупных человекообразных обезьян . Уже у черепа Sangiran 2 большое затылочное отверстие сдвинуто вперёд — анатомический признак сформировавшихся навыков прямохождения . У яванских Homo erectus классического периода свод черепа выраженной грушевидной формы, сужающийся ниже глазниц. Надбровные валики неизменно массивные, высокие и соединённые по центру ( глабелла не только не вдавлена, но даже выступает вперёд ), они всегда отделены от чешуй височных костей треугольными впадинами. Напротив, височные валики могут существенно различаться по степени выраженности. Сосцевидный отросток височной кости (направленный вниз отросток височной кости, расположенный позади наружного слухового прохода, с воздухоносными ячейками), заметно выдающийся за основание черепа у наиболее ранних экземпляров, у более поздних может быть выражен в значительно меньшей степени. Лицевой отдел широкий, челюсти умеренно выдаются вперёд. Найдено множество отдельных нижних челюстей, но они настолько разнообразны, что часть исследователей считает их принадлежащими разным биологическим видам. В целом челюсти классического периода достаточно высокие для своей ширины, что может быть связано с большой жевательной поверхностью моляров . Нижние резцы широкие, моляры отличают высокие режущие кромки . Длина моляров уменьшается по мере удаления от центра. Клыки более крупные, чем у современного человека .
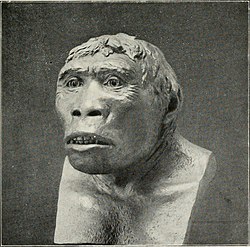
Черепа индонезийских и китайских представителей вида H. erectus в целом схожи по форме и размеру, их характеризует длинный и низкий свод. В отличие от индонезийских, однако, китайские черепа сужаются в районе астерионов — симметричных точек в нижней части затылка, а также фронтально, в предглазничной области. Кроме того, черепа синантропов отличают меньшая толщина (у яванских черепов кости черепа очень толстые даже у находок с поздними датировками ) и менее выраженный изгиб надглазничных валиков (у африканских представителей вида валики ещё меньше) . Яванские черепа по сравнению с африканскими и китайскими характеризуются очень узкой лобной костью и (в случае находок из Сангирана) гипертрофированно широкой затылочной . Почти полное отсутствие теменной вырезки в височной кости рассматривается как признак специализации вследствие длительной островной изоляции . У питекантропа заметно более выпуклый лицевой отдел и, по-видимому, (с учётом погрешностей реконструкции) более выраженный прогнатизм , чем у синантропа и африканского H. erectus , более высокие челюсти и в целом более крупные зубы . Челюсти питекантропов настолько массивны и обезьяноподобны, что при первых их находках учёные не смогли отождествить их с обладателем прогрессивной бедренной кости и описали как остатки нового вида Meganthropus palaeojavanicus . Только с обнаружением более полных черепов с лицевым отделом (в том числе Sangiran 17 и Sangiran IX) стала окончательно ясна принадлежность челюстей, за которыми, однако, и после этого в научной терминологии сохранилось название «мегантропные» .
Не считая черепов и челюстей, детали скелета человека прямоходящего достаточно фрагментарны, и во многих случаях существуют сомнения по поводу видовой принадлежности. Тем не менее известно, что, как и у других представителей рода Homo , у длинных костей человека прямоходящего были бо́льшая, нежели у австралопитеков , площадь суставных поверхностей, более толстая трубчатая часть, двойное менисковое соединение проксимальной стороны большеберцовой кости , узкие подвздошные кости с чётко выраженными столбами (включая ацетабулярно-гребешковую подпорку) и медиальное осевое вращение седалищного бугра . Кроме того, человек прямоходящий крупнее не только австралопитеков, но и человека умелого — более древнего представителя рода Homo . По сравнению с этими гомининами его верхние конечности короче относительно нижних, а верхнее открытие таза, по-видимому, шире, что позволяет рождение детей с бо́льшей головой. Наряду с прогрессивными чертами скелет Homo erectus обладает и рядом примитивных характеристик, не встречающихся у современных людей. В их число входят длинные шейки бедренных костей и шесть поясничных позвонков .
Вместе с костями питекантропа и других представителей вида Homo erectus часто обнаруживаются кости других животных и окаменелые остатки растений. Эти окаменелости, а также специфика пород, в которых они обнаруживаются, дают основу для предположений о том, в каких условиях существовал человек прямоходящий .
В частности, почвы в районе Сангиранского холма указывают на существование в этот период пресноводных озёр (верхняя Сангиранская формация) и разветвлённой системы ручьёв (Бапангская формация, названная в честь близлежащей деревни ). Отложения раннего плейстоцена — как органический материал в целом, так и остатки пресноводных моллюсков в частности — характеризует высокий показатель δ 13 C . Этот параметр свидетельствует об обилии растительности типа C 4 . Если в более умеренном климате такая растительность означала бы наличие саванн , то в приэкваториальных широтах растения этого типа предпочитают влажные почвы. В целом отложения данного слоя характерны для низменной местности с заболачиваемыми лугами и обилием озёр, вероятно, в верховьях мелководного эстуария . По-видимому, к моменту появления первых людей в этих краях там произрастали папоротники , осоковые , другие водолюбивые травы и деревья и обитали животные, ведущие полуводный образ жизни (небольшие гиппопотамы , крокодилы ) и различные оленевые .
В Бапангской формации (более недавний слой почв, залегающий над почвами Сангиранской формации) происходит постепенное смещение значений δ 13 C, указывающее на замещение трав типа C 4 растительностью типа C 3 , характерной для более сухого климата. На то, что климат в регионе со временем становился более сухим, указывают и меняющаяся корневая структура, и уровни накопления карбонатов в почве, повышающиеся из-за сезонного недостатка влаги. Около 1,3 миллиона лет назад регион, по-видимому, покрывала разветвлённая система ручьёв и рек, через которые уходила вода со склонов близлежащих вулканических нагорий. Местность представляла собой открытую лесостепь — саванну, перемежаемую редкими рощами и полосами деревьев вдоль речных русел. Животный мир был представлен стегодонами и различными представителями свиных и бычьих , найдены также кости карликовых бегемотов , гульманов Semnopithecus auratus и макаков-крабоедов . Аналогичные отложения характерны также для Триниля, где, помимо этого, встречались крупный эндемичный тигр и бенгальская кошка , несколько видов обезьян, а из крупных травоядных — яванский носорог .
Ещё позднее, на протяжении отрезка между 900 и 130 тысячами лет назад, как минимум часть которого совпадает с находками костей классического питекантропа, климат и природные условия продолжали меняться . Эрозия окружающих возвышенностей привела к накоплению в районе Сангирана твёрдого осадочного слоя толщиной от 1 до 4 метров, состоявшего из песчаника и вулканических пород и известного как grenzbank (гринзбанк ). Происходили значительные изменения в животном мире: архаичные виды фауны, населявшие открытую лесостепь, вымирали, уступая место фауне современных тропических дождевых лесов , известной в палеонтологии как пунунгская. По всей видимости, к моменту полного исчезновения архаичной сангиранской и нгандонской фауны и замещения её пунунгской питекантропов на Яве уже не оставалось . Слой гринзбанка в сангиранских отложениях разделяет два отличных типа фоссилий человека прямоходящего — архаичный или классический тип (питекантроп) ниже него и более прогрессивный (явантроп) выше .
В целом в комплексах ископаемых остатков, найденных на Яве, заметно преобладают остатки травоядных животных (для сравнения — среди остатков плейстоценовой фауны в смоляных ямах Ранчо Ла-Брея больше костей хищников) . При раскопках в Триниле, Кедунг-Брубусе и ряде других локаций многочисленные кости крупных травоядных (преимущественно бычьих, а также оленьих и хоботных ) соседствуют со значительно более редкими костями питекантропа. Редкость человеческих остатков, сопоставимых по частоте с остатками крупных хищников, а не травоядных, позволяет выдвинуть предположение, что и в рационе питекантропов на этом этапе развития значительную долю составляло мясо, поскольку для хищников характерна меньшая плотность расселения . Включение в диету мяса с его более высокой, чем у растительной пищи, энергетической ценностью могло обуславливаться увеличением общих размеров тела человека (в особенности головного мозга) и большей энергозатратностью передвижения на двух ногах .
Потребность в мясе, в свою очередь, должна была способствовать как использованию рукотворных орудий, применяемых при охоте , так и более быстрому расселению в поисках добычи . В то же время анализ богатого культурного слоя на стоянках человека прямоходящего в Африке позволяет утверждать, что человеческая деятельность на этих стоянках продолжалась долгое время. Это могло быть результатом либо оседлого образа жизни, либо цикличных миграций с периодическими возвращениями на прежнее место, подобно тому, как это зафиксировано у аборигенов Австралии и других современных племён охотников-собирателей .
Фоссилии человека прямоходящего предоставляют определённую информацию о свойственных ему патологиях . Так, в литературе обращается особое внимание уже на первую бедренную кость питекантропа, найденную Дюбуа: она демонстрирует наличие экзостоза , или патологического формирования новой костной ткани на поверхности уже существующей кости . Предполагается также, что изменения анатомии человека при переходе от Homo habilis к Homo erectus повлекли за собой изменения и в социальных отношениях в стаде . Увеличившийся объём головы младенца означал, что матери требовалась посторонняя помощь при родах и в восстановительный период после них. Согласно существующим теориям, из-за появления таких помощников, заботящихся о матери и младенце в первое время после родов, шансы новорождённых на выживание выросли . В то же время высказывалось мнение, что адаптация к прямохождению шла медленно, и из-за многочисленных осложнений при родах смертность среди рожениц была высокой. Это, в свою очередь, должно было приводить к стабильному численному перевесу самцов среди взрослых особей в стаде людей прямоходящих .
Если находки фоссилий классического человека прямоходящего на Яве достаточно многочисленны, этого нельзя сказать о предметах материального быта. В целом в мире обнаружены многочисленные примитивные орудия, ассоциируемые с человеком прямоходящим эпохи нижнего и среднего плейстоцена и относимые к так называемой ашельской культуре . Сообщения о находках орудий труда питекантропа на Яве появлялись начиная с 1911 года, когда было объявлено об обнаружении в том же горизонте, что и тринильские окаменелости, костей животных со следами обработки. В 1930-х годах фон Кёнигсвальд сообщил о находке маленьких каменных орудий в Сангиране, месте обнаружения основной массы костей древнего человека на Яве. В этом же десятилетии в южной части острова, в , были найдены более крупные каменные инструменты с более заметными следами обработки, также первоначально приписанные питекантропу. Наконец, в 1970-е годы каменные орудия ( чоппер и отщеп ) были найдены в Самбунгмачане — между Тринилем и Сангираном. Эти орудия публикация 1978 года датировала средним плейстоценом .
Впоследствии, однако, было показано, что следы на костяных «орудиях» из Триниля, скорее всего, оставлены зубами хищников и падальщиков , а каменные орудия из Сангирана и Пачитана относятся к верхнему плейстоцену и (в последнем случае) даже голоцену и, соответственно, не могут быть частью материальной культуры классического человека прямоходящего. Находки из Самбунгмачана, обнаруженные неподалёку от костей явантропов, напоминают аналогичные орудия из более поздних слоёв и поэтому, возможно, переотложены и на самом деле созданы не в среднем, а в верхнем плейстоцене .
В свете переоценки ранних находок было высказано мнение, что при поиске орудий питекантропа следует сосредоточиться не на чопперах и подобных инструментах (ассоциируемых с более поздней формой H. erectus ), а на грубых отщепах и камнях с минимальными следами обработки . Новые находки каменных орудий именно такого типа, датируемых периодом между 800 тысячами и 1,51 миллиона лет назад, были сделаны в конце XX — начале XXI века при раскопках в Сангиране. Исследовательская группа Гарри Видианто раскопала несколько сотен примитивных отщепов длиной несколько сантиметров каждый, преимущественно из халцедона . В то же время, хотя большинство отщепов были обкатаны водой, ряд инструментов, выполненных из песчаника, сохранили режущие кромки, что может означать, что и в данном случае произошло переотложение из более поздних слоёв . Ещё одна находка, сделанная в археологической локации Нгебунг в слоях возрастом около 800 тысяч лет, включает более разнообразные орудия, в том числе чоппер, рубило и большой режущий инструмент. Для этих орудий тоже характерна примитивная обработка, лишённая прогрессивных черт, отличающих орудия из Африки и Индии .
Возможно также, что редкость каменных орудий эпохи раннего палеолита на Яве связана с тем, что питекантропы использовали в основном не камень, а менее долговечные (но более лёгкие в обработке) материалы — например, кость и дерево, в том числе бамбук . Подтверждением этой гипотезы могут служить находки, сделанные ещё самим Дюбуа. Вместе с первыми костями питекантропа нидерландский естествоиспытатель собрал значительное количество раковин моллюсков рода Pseudodon , возраст которых при позднейшем радиоизотопном датировании был оценён в 380—640 тысяч лет. На некоторых из них были обнаружены отверстия диаметром 0,5—1 см в месте, где у живого моллюска располагается мускул-замыкатель. На ряде обнаруженных вместе с раковинами акульих зубов были найдены следы износа, что позволяет предположить, что их использовали для высверливания дырок в раковинах. Кроме того, на одной из раковин имелись следы износа, указывающие, что ею пользовались как режущим и скоблящим инструментом . В Сангиране найдены также датируемые периодом 1—1,6 миллиона лет назад кости крупных травоядных со следами надрезов, форма которых отличается от надрезов, которые делаются каменными орудиями, но соответствует следам от кромок раковин моллюсков .

На одной из раковин из Триниля были обнаружены геометрические узоры в форме букв «M» и «N», по-видимому, процарапанные зубом акулы. Сообщившие о находке исследователи полагают, что царапины нанесены не случайным образом, а с целью получения непрерывного узора, и рассматривают эту резьбу как древнейший образец искусства архаичного человека, как минимум на 300 тысяч лет более ранний, чем другие известные гравировки .
Большинство специалистов, опираясь на археологические свидетельства из Китая, сходится во мнении, что древний человек начал использовать огонь не менее чем 0,5 миллиона лет назад. Однако в отличие от Китая, где известны многочисленные следы использования огня в сочетании с фоссилиями синантропа, на Яве обожжённое дерево в соответствующих слоях до конца 1980-х годов было обнаружено только один раз. Данная находка была сделана в Триниле. Точная датировка находки затруднена и охватывает период от 0,5 до 0,83 миллиона лет назад, но авторы публикаций, посвящённых ей, указывают на то, что данный регион вулканически активен и огонь мог иметь естественное происхождение. Таким образом, нет однозначных доказательств, что питекантропы умели добывать огонь или хотя бы использовать огонь естественного происхождения для своих нужд . Если даже подобное ситуативное использование природного огня имело место, его невозможно подтвердить на основе доступного археологического материала .
|
|
|
|---|---|
| Таксономия |
|
|
Вымершие роды
Гоминини / Хоминина |
|
|---|---|
| Люди (род Homo ) | |
| Находки гоминид | |
| Происхождение |
|
| Распространение | |
|
|
Эта статья входит в число
избранных статей
русскоязычного раздела Википедии.
|