Interested Article - Эдмонтозавр

- 2021-09-06
- 1
Эдмонтоза́вр ( лат. Edmontosaurus , переводится как «ящер из Эдмонта», от названия города Эдмонтон в Канаде и др.-греч. σαύρα — ящер) — род динозавров из семейства гадрозаврид (утконосых динозавров) , который относят к гадрозавринам (зауролофинам) — подсемейству гадрозавридов, не имевших гребня. Окаменелости этих древних пресмыкающихся были обнаружены в западной части Северной Америки в отложениях возрастом от 73 до 66 млн лет . Ископаемые остатки эдмонтозавров многочисленны и хорошо изучены, обнаружено даже несколько окаменевших мумий .
Род был впервые описан в 1917 году по остаткам, обнаруженным на юге Канады , однако отдельные особи описывались как до этого, так и позднее как представители других родов: Claosaurus , Thespesius , Trachodon и Anatosaurus . Эдмонтозавры были одними из крупнейших гадрозавридов, до 15 м в длину и массой до 3,2 т , и были широко распространены в Северной Америке (окаменелости обнаруживались как в Колорадо , так и на Аляске ). Изучение пород , в которых были найдены кости эдмонтозавров, позволяет предположить, что они населяли прибрежные равнины. Эдмонтозавры были растительноядными, передвигались в основном на четырёх ногах, но при необходимости могли подниматься на задние. Известные остатки со следами зубов позволяют предположить, что эдмонтозавры могли быть объектами охоты тираннозавров , а массовые захоронения в костеносных слоях — что они были стадными животными .
Описание

Как и другие гадрозавриды , эдмонтозавры были крупными животными с длинным, уплощённым с боков хвостом и с широким «утиным клювом» на конце морды. Передние ноги эдмонтозавров были не такими длинными и мощными, как задние, но достаточно длинными, чтобы помогать при ходьбе или в стоячем положении. В отличие от многих других гадрозавридов, череп эдмонтозавров был лишён гребня — полого или сплошного; некое подобие петушиного гребня при жизни животного могли образовывать мягкие ткани . Эдмонтозавры относятся к крупнейшим гадрозавридам — взрослые особи наиболее крупных видов могли достигать в длину от 9 до 12 , 13 или даже 15 м , а весили от 2,5 до 3,2 т (эта оценка может быть значительно увеличена, если подтвердится теория, причисляющая к эдмонтозаврам гигантского азиатского ящера шантунгозавра , масса тела которого оценивается в 16 т ). Традиционно крупнейшим видом эдмонтозавров считается Edmontosaurus regalis (голотип вида, NMC 2288, достигает не менее 9 м в длину ), хотя существует гипотеза, согласно которой более крупный гадрозаврид Anatotitan copei является на самом деле представителем вида Edmontosaurus annectens . Эту гипотезу выдвинули в 2004 году Дж. Хорнер с соавторами , а в конце первого и начале второго десятилетия XXI века в её поддержку выступили Н. Кампион и Д. Эванс . Если не рассматривать теорию Хорнера, Edmontosaurus annectens считается менее крупным видом: длина двух наиболее известных полных скелетов, USNM 2414 и YPM 2182, составляет соответственно 8 и 8,9 м . Однако есть вероятность, что речь идёт о подростковых особях , и в 1970 году сообщалось о намного более крупном скелете (почти 12 м в длину), потенциально принадлежащем Edmontosaurus annectens , в коллекции Лос-Анджелесского окружного музея . Ещё один выделявшийся ранее вид, Edmontosaurus saskatchewanensis (в настоящее время рассматривается в рамках вида Edmontosaurus annectens ), был меньше, примерно 7 м длиной .
Строение скелета

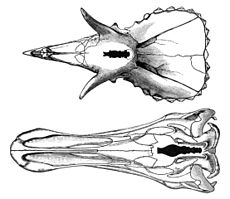
Длина черепа взрослого эдмонтозавра могла превышать метр. Один из известных черепов E. annectens (ранее классифицировавшегося как анатотитан) достигает в длину 1,18 м при общей длине скелета 11,5 м . В профиль череп эдмонтозавра имел приблизительно треугольную форму , костяной гребень отсутствовал . Если смотреть на череп сверху, то можно заметить, что его «клюв» был широким, как у утки или колпицы . Клюв был покрыт слоем кератина и не имел зубов — последняя черта характерна для всех гадрозавридов, кроме примитивного тельматозавра . Значительная часть такого кератинового покрытия сохранилась на верхней части клюва «мумии» из коллекции Зенкенбергского музея естественной истории . У этого экземпляра сохранившаяся кератиновая часть клюва выдаётся вперёд по отношению к кости не менее чем на 8 см и направлена отвесно вниз по отношению к ней . У экземпляра из Лос-Анджелесского окружного музея клюв с верхней челюсти выдаётся вперёд на 11 см и опускается на 5 см ниже нижней челюсти , перекрывая её переднюю поверхность .

Зубы присутствовали на верхней и нижней челюстной кости . Зубы эдмонтозавра (как и других гадрозавридов) соединялись в так называемые «батареи», в каждой из которых было до шести зубов . У разных видов эдмонтозавра имелось разное количество зубных колонн в батарее: 51—53 на верхней челюсти и 48—49 на нижней у E. regalis , 43 на верхней и 36 на нижней у E. annectens и 52 на верхней и 44 на нижней у E. saskatchewanensis . Зубы непрерывно сменялись, формирование новых зубов занимало от 1/2 до 2/3 года . В их конструкции были задействованы шесть видов тканей, что делает их сравнимыми по сложности с зубами млекопитающих, причём взаимное расположение тканей в зубе могло изменяться по мере изменения его положения в батарее и во рту (так, эмаль покрывала только внешнюю сторону зубов на верхней челюсти и внутреннюю — на нижней ).
Носовые отверстия эдмонтозавра имели форму вытянутых овалов и располагались в глубоких впадинах, окружённых сверху, сзади и снизу ярко выраженными костяными каёмками . Заглазничная кость образует «карман», встречающийся и у других гадрозавридов, но наиболее ярко выраженный и глубокий именно у эдмонтозавров . По крайней мере в одном случае (у экземпляра в Зенкенбергском музее) в глазных впадинах сохранились склеротикальные кольца (маленькие косточки, которые поддерживают глазное яблоко ) — редкий в палеонтологии случай, позволяющий более надёжно определить положение глаз . У представителей одного из видов эдмонтозавров было найдено стремечко — маленькая кость среднего уха, также редко сохраняющаяся у ископаемых .
Число позвонков отличается у разных видов. У E. regalis было 13 шейных, 18 спинных, 9 крестцовых и не менее 29 хвостовых позвонков . У одного экземпляра, первоначально отнесённого к виду Anatosaurus edmontoni (ныне причислен к виду E. regalis ), был обнаружен дополнительный крестцовый позвонок, а число хвостовых оценивается в 85, но эта оценка в большой степени предположительна . Для гадрозавридов как семейства число хвостовых позвонков варьирует от 50 до 70 , поэтому оценка количества хвостовых позвонков этого экземпляра, возможно, является завышенной.
При эдмонтозавр держал грудную часть туловища наклонённой вниз, а шею, вероятно, держал поднятой вверх, в результате чего передняя часть его позвоночника выгибалась U-образно. Хвост и поясничную область эдмонтозавр держал параллельно поверхности земли . Вдоль большей части спины и хвоста проходила сетка оссифицированных сухожилий, отходивших от позвоночника. Такое строение делало спину и по крайней мере часть хвоста, по выражению палеонтолога Питера Галтона, «прямыми, как палка» . Предполагается, что оссифицированные сухожилия помогали расположенному в основном горизонтально позвоночному столбу поддерживать тяжесть огромного тела, опиравшегося лишь на задние ноги .

Таз эдмонтозавра состоял из трёх элементов:
- вытянутой подвздошной кости над местом соединения с ногой;
- седалищной кости с длинной тонкой ветвью, расположенной ниже и сзади;
- и расширяющейся наподобие тарелки лобковой кости впереди.
Из-за строения бедра эдмонтозавр не мог принимать вертикальное положение, так как в таком положении бедренная кость просто выскочила бы из тазобедренного сустава. Таз поддерживал соответствующий отдел позвоночника, состоявший из девяти слитых позвонков. Бедренная кость эдмонтозавра была прямой и мощной, примерно посередине её длины имелся выступ для прикрепления мышц . Этот выступ служил местом крепления мощных мышц бёдер и хвоста, оттягивающих задние ноги ещё больше назад и помогающих использовать хвост для поддержания равновесия . На каждой задней ноге имелось по три пальца, среди которых отсутствовали большой и мизинец. На всех трёх пальцах имелись копытообразные ногти .
Лопатки эдмонтозавра были длинными и плоскими, располагались они почти параллельно линии позвоночного столба . Передние конечности были менее длинными и мощными по сравнению с задними. На плечевой кости имелся развитый дельтопекторальный узел для прикрепления мышцы. Локтевая и лучевая кости были относительно утончёнными и хрупкими, плечевая кость и предплечье были примерно одной длины. Запястье было устроено довольно просто, включая всего две кости. На каждой передней конечности было четыре пальца, ни один из которых не был противостоящим (при первых описаниях была допущена ошибка, и некоторое время в этом качестве фигурировал короткий и толстый мизинец). Третий и четвёртый пальцы были приблизительно одинаковой длины и при жизни были сращены . Пальцы передней конечности при жизни покрывала общая кожаная подушка, но третий палец был снабжён копытообразным ногтем, обеспечивавшим дополнительную опору при ходьбе на четырёх ногах .
Кожный покров


На многих экземплярах E. annectens были обнаружены остатки кожи. Некоторые из таких экземпляров широко известны — среди них «мумия траходона», подробно описанная в начале XX века , и найденный век спустя экземпляр, получивший прозвище «Дакота» , у которого сохранились даже органические вещества кожного покрова . Палеонтологи обнаружили «Дакоту» в 2004 году на территории штата Северная Дакота в США. Исследователи два года удаляли породы, в которых была заключена находка, после чего их глазам предстали окаменелая кожа, связки, сухожилия . Благодаря этим находкам можно практически полностью восстановить текстуру шкуры для вида E. annectens . Сохранившихся отпечатков кожи E. regalis известно меньше, но существуют описания таких экземпляров, включая публикацию 2013 года, сообщающую об особи с гребнем из мягких тканей наподобие петушиного. Эта находка породила гипотезы о социально-иерархическом или социально-половом значении таких гребней и их возможном существовании у других динозавров .
«Мумия траходона» — экземпляр из Американского музея естественной истории , известный как AMNH 5060, — представляет собой окаменевшую естественную мумию и в настоящее время классифицируется как представитель вида E. annectens . У этого экземпляра видны остатки кожи на морде, большой части шеи и туловища и частично на кистях и ногах . Хвост и часть ног не сохранились и структура кожного покрова для этих частей тела экземпляра неизвестна, а верхние слои эпидермиса вокруг шеи и на передних конечностях были случайно повреждены в начале препарирования окаменелостей . Предполагается, что данная особь мумифицировалась в сухом речном русле . Обстоятельства местоположения и позы тела позволяют предположить, что животное умерло во время длительной засухи, возможно, от истощения . Впоследствии мумифицированный труп был занесён грунтом в ходе внезапного паводка, на что указывают отложения, в которых он найден: они содержали достаточно частичек глины, чтобы сделать возможным отпечаток кожных покровов .
Как показывают исследования «мумии траходона», его эпидермис был очень тонким для животного подобных размеров , чешуйки не перекрывались друг с другом и напоминали чешуйки ящерицы жилатье . На большей части тела было только два вида чешуек:
- «Грунт» ( англ. ground plan ) — маленькие и выпуклые или заострённые (бугорки) в диаметре 1—3 мм , без определённого порядка расположения;
- «Тротуарная плитка» ( англ. pavement ) — плоские и многоугольные чешуи большего размера, но обычно не более 5 мм в диаметре, только на предплечье достигавшие 10 мм в диаметре. Чешуи второго типа располагались группами, которые разделялись участками кожи с чешуёй первого типа, причём на стыках встречаются чешуи переходной формы. На большей части тела чешуйки были сгруппированы в круглые или овальные участки, но на плече чешуйки группировались рядами, параллельными лопатке. Вообще, группы чешуек были большими на верхней части тела (над бёдрами достигая в длину 50 см) и мелкими на нижней стороне .

Отпечатки чешуи на голове находятся в области больших отверстий, где располагались ноздри. В этой части вместо чешуй сохранился отпечаток мягких кожных складок, более глубоких в передней части отверстий — в месте, где была собственно сама ноздря . Вдоль шеи и спины тянулся мягкий гребень или кожная складка, а над позвонками располагалась цепочка чешуйчатых участков, состоящих из туберкул овальной формы. Общая высота гребня у AMNH 5060 неизвестна, как и то, где находился его верхний конец, поскольку в этом районе отпечаток был утерян при препарировании, но гребень был не меньше 8 см в высоту и лежал складками, что облегчало движение. Палеонтолог Генри Осборн , первым описавший эту мумию, высказывал предположение, что высота гребня была достаточной, чтобы на нём разместился ещё как минимум один ряд чешуйчатых участков .
На предплечьях «мумии траходона» располагаются самые крупные чешуи, сгруппированные на больших участках и покрывающие переднюю поверхность конечности. Кисти были покрыты мелкими бугорками первого типа, покрывающими мягкую ткань, в которую заключены три центральных пальца, включая ногти. Осборн считал, что эта конструкция представляла собой плавательную перепонку , но позже Роберт Баккер пришёл к выводу, что это была подушечка из мягких тканей, помогавшая при ходьбе, подобно существующим у верблюдов — наличие такой подушечки подтвердилось, когда была обнаружена мумия «Дакота» . Как и на предплечье, на голени чешуйки были крупными; какая чешуя покрывала остальную часть ног эдмонтозавра, неизвестно, хотя известные отпечатки у другого гадрозаврида — ламбеозавра — показывают, что кожа с тела могла покрывать ляжки, как у современных птиц .
Хотя хвост AMNH 5060 не сохранился, известны отпечатки кожи хвоста у некоторых других экземпляров. Отпечатки кожи с частично сохранившегося хвоста E. annectens , обнаруженного в формации Хелл-Крик (Монтана), показывают наличие сегментированного гребня над позвоночником. У этого экземпляра спинной гребень достигал 8 см в высоту, сегменты достигали около 5 см в ширину и 4,5 см в толщину (в продольном сечении) и располагались на расстоянии 1 см один от другого, по одному сегменту на позвонок . Другой хвост, как удалось определить, принадлежал неполовозрелой особи вида Е. annectens ; на этой окаменелости сохранились как отпечатки уже известных туберкул, так и до этого не встречавшихся кожных покровов: накладывающихся чешуй эллиптической формы, чешуй с желобками и «трапециевидной рогоподобной структуры размерами 9 на 10 см » .
Мозг и нервная система
Описания мозга эдмонтозавра, фигурирующие в ряде исследовательских публикаций, основываются на анализе эндокранов внутренней полости черепной коробки известных экземпляров. В их число входят описания мозга E. annectens и E. regalis , а также экземпляров эдмонтозавра, видовая принадлежность которых не установлена . По сравнению с размерами тела эдмонтозавра его мозг был небольшим — длина содержавшей его полости составляла только четверть от общей длины черепа , и по различным оценкам её объём составлял от 374 до 450 см³ , из которых мозг мог занимать лишь немногим более половины , а остальное пространство заполняла поддерживающая его твёрдая мозговая оболочка . Так, объём мозга у особи с объёмом полости 374 см³ , по оценкам Г. Джерисона, Дж. Хорнера и С. Хорнера, составлял только 268 см³ . Мозг представлял собой продолговатую структуру , в которой, как и у других позвоночных за исключением млекопитающих, отсутствовала новая кора . Хорошо развитые структуры представляли собой эпифиз и гипофиз . Как и у стегозавров , нервная трубка у эдмонтозавров утолщалась в районе крестца, но не в такой же степени: если у стегозавров объём, занимаемый этим «задним мозгом», в 20 раз превосходил объём мозговой полости, то у эдмонтозавров это соотношение было всего лишь 2,59:1 .
Систематика
| Кладограммы гадрозавридов (зауролофидов) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эдмонтозавры относились к гадрозавридам — утконосым ящерам, семейству динозавров, к настоящему времени известному только по ископаемым остаткам из верхнемеловых отложений. В рамках семейства гадрозавридов эдмонтозавры принадлежат к подсемейству гадрозаврин (с 2010 года в части источников называемых зауролофинами ), отличавшемуся от ламбеозаврин отсутствием полого головного гребня. Другими представителями этого подсемейства были брахилофозавр , майазавр , грипозавр , прозауролоф , наашойбитозавр , зауролоф , критозавр и лофоротон . Очень похож на эдмонтозавров крупный гадрозаврид из Китая — шантунгозавр ; палеонтолог Майкл Бретт-Серман утверждает, что они ничем не отличаются кроме того, что приписываемые шантунгозавру остатки были более крупными . Описанный в 2016 году по фоссилиям неполовозрелых особей вид зауролофин в более поздних публикациях также расценивается как nomen dubium , а лёгший в основу его описания материал — как принадлежащий очень молодым эдмонтозаврам неопределённого вида .
Хотя принадлежность эдмонтозавров к гадрозавринам не оспаривается, точное место эдмонтозавра на кладограмме гадрозавридов не установлено. На раннем этапе развития филогенетики , в частности в монографии и Н. Райт, вышедшей в 1942 году, эдмонтозавры и анатозавры (большинство видов которых в дальнейшем будут также отнесены к эдмонтозаврам) были представлены как одна из ветвей «гладкоголовых» гадрозавров . Один из первых кладистических анализов связал эдмонтозавров с анатотитаном и шантунгозавром в неформальную кладу эдмонтозавров, наиболее близкую к зауролофам и состоящую в более удалённом родстве с брахилофозаврами и грипозаврами . Те же результаты получили Терри Гейтс и Скотт Сампсон в 2007 году . В 2004 году Дж. Хорнер предположил, что эдмонтозавр был близким родственником майазавра и брахилофозавра, а зауролоф, наоборот, по эволюционному родству отстоит от него дальше , но ряд филогенетических исследований второго десятилетия XXI века снова продемонстрировал, что родство эдмонтозавра с майазавром и брахилофозавром более отдалённое, а его ближайшими родичами являются шантунгозавр и зауролофы .
История изучения
Claosaurus annectens и другие ранние находки


Почти полвека видовая классификация эдмонтозавров была крайне запутанной, многие экземпляры выделялись в отдельные рода. На разных этапах истории их ассоциировали с такими родами, как агатаумы , анатозавры, анатотитаны, клаозавры , гадрозавры , теспесии и траходоны (в настоящее время некоторые из этих родов не рассматриваются как отдельные таксоны). До 1980-х годов, в зависимости от авторов и времени публикаций, окаменелые остатки эдмонтозавров, за исключением вида E. regalis , классифицировались как ископаемые остатки анатозавра, клаозавра, теспесия или траходона.
Хотя само родовое название Edmontosaurus появилось лишь в 1917 году, E. annectens — первый из хорошо изученных видов, в настоящее время относимых к этому роду — был описан уже в 1892 году как вид клаозавров. Согласно справочнику The Dinosauria, первое описание окаменелостей, возможно, принадлежащих эдмонтозаврам, было сделано уже в 1871 году, когда американский натуралист Эдвард Коп описал новый вид Trachodon cavatus (в более поздних источниках название передаётся как T. avatus и даже T. atavus ). Впоследствии данные остатки причислили к описанному в 1917 году E. regalis . В 1874 году Коп дал название ещё одному новому виду Agathaumas milo ; этот таксон был выделен (но не описан) по крестцовым позвонкам и фрагментам берцовых костей маастрихтской (верхний мел) в Колорадо . В этом же году он отнёс эти остатки к виду Hadrosaurus occidentalis . До настоящего времени эти остатки не сохранились . Как и Trachodon cavatus , Agathaumas milo — видовое название, предшествующее Edmontosaurus regalis по времени — рассматривается в позднейших обзорах как его синоним . Ни первый, ни второй из описанных Копом видов не привлекли широкого внимания — в частности, они даже не упоминаются в монографии Лалла и Райт 1942 года. Третий из ранних сомнительных видов, Trachodon selwyni , описанный Лоуренсом Ламбом в 1902 году по нижней челюсти из формации в Альберте (Канада), известной сейчас как , упоминается Лаллом и Райт как «сомнительный» (в 1997 году ошибочно сообщалось, что Лалл и Райт рассматривали Trachodon selwyni как эдмонтозавра ). Авторы новых обзоров сходятся в своих оценках T. selwyni с Лаллом и Райт .
Первая хорошо документированная окаменелость, в настоящее время классифицируемая как эдмонтозавр, была описана в 1892 году Г. Ч. Маршем под названием Claosaurus annectens . У голотипа этого вида, известного под номером USNM 2414, частично сохранились черепная крышка и скелет; при описании использовались также второй череп и скелет, принадлежащие экземпляру YPM 2182, который рассматривается как паратип данного вида. Оба экземпляра были найдены Дж. Б. Хетчером в позднемаастрихтской (Вайоминг) . Claosaurus annectens стал одним из первых видов динозавров, для которого была проведена реконструкция скелета, и первым гадрозавридом, для которого такая реконструкция была признана успешной . YPM 2182 и UNSM 2414 являются соответственно первым и вторым относительно полными восстановленными и выставленными на обозрение скелетами динозавров на территории США: экземпляр YPM 2182 был выставлен в музее в 1901 году, а USNM 2414 в 1903 году .
Из-за неполноты представлений о гадрозавридах в этот период Claosaurus annectens после смерти Марша в 1897 году попеременно классифицировался как вид клаозавров, теспесиев или траходонов. Расхождения во мнениях были очень заметными: учебники и энциклопедии иллюстрировали различия между «игуанодоновидным» Claosaurus annectens и «утконосым» гадрозавром, пользуясь ископаемыми остатками (как сейчас установлено) взрослых особей Edmontosaurus annectens , и в то же время Хетчер писал о C. annectens как о гадрозавриде, ссылаясь на те же самые «утконосые» черепа . В публикации 1902 года Хетчер занял радикальную позицию, определив почти все принятые на тот момент родовые названия гадрозавридов как синонимы рода Trachodon . В число этих «траходонов» попали , , Hadrosaurus и Ornithotarsus , и Thespesius , а также и , в настоящее время считающиеся рогатыми динозаврами . На основе работ Хетчера в палеонтологическом сообществе был достигнут кратковременный консенсус, пока во втором десятилетии XX века новые находки из Канады и Монтаны не показали, что гадрозавриды более разнообразны, чем считалось до этого . В 1915 году Чарльз Гилмор пересмотрел классификацию гадрозавридов, сделав вывод, что для окаменелостей Лансской формации и современных им должно быть возвращено родовое имя Thespesius , а название Trachodon , ввиду нехватки материала, следует использовать только для окаменелостей более раннего периода ( и её эквиваленты). Claosaurus annectens Гилмор рекомендовал считать синонимом Thespesius occidentalis .
В период между 1902 и 1915 годами были сделаны ещё две важные находки особей Claosaurus annectens . Первая, AMNH 5060, известная как «мумия траходона», была обнаружена в 1908 году Чарльзом Штернбергом и его сыновьями в отложениях Лансской формации недалеко от Ласка (Вайоминг). Находка была сделана, когда Штернберг работал на лондонский Музей естествознания , но в итоге была продана за 2000 долларов Г. Ф. Осборну в Американский музей естественной истории . Ещё один похожий экземпляр был найден Штернбергами в том же районе в 1910 году ; у этого экземпляра также сохранились, хотя хуже, чем у первого, отпечатки кожи. Этот экземпляр (SM 4036) был продан Зенкенбергскому музею в Германии .
Канадские находки
В 1917 году Лоуренс Ламб впервые ввёл родовое имя Edmontosaurus , опираясь на два неполных скелета из нижней Эдмонтонской формации (ныне ) на реке Ред-Дир на юге канадской провинции Альберта . Новый род получил название в честь Эдмонтонской формации , отложения которой старше тех, в которых был впервые обнаружен Claosaurus annectens . Типовой вид , E. regalis («королевский» или, в переносном смысле, «очень большой») , описан по экземпляру NMC 2288, состоящему из черепа, хорошо сохранившегося позвоночника до шестого хвостового позвонка, рёбер, частично сохранившихся костей таза и задней конечности и одной из плечевых костей ; этот экземпляр был найден в 1912 году Леви Штернбергом. От второй особи, паратипа NMC 2289, сохранились череп и скелет за исключением «клюва», части хвоста начиная с пятого хвостового позвонка и части ступней. Она была обнаружена в 1916 году ещё одним сыном Чарльза Штернберга — Джорджем. Ламб отметил, что новый вид имеет значительное сходство с Diclonius mirabilis (известным по особи, которая в настоящее время считается представителем вида Edmontosaurus annectens ), но подчеркнул общую величину, мощное сложение и более высокий череп эдмонтозавра . Первоначальное описание в основном фокусировалось на черепах обеих особей, но в 1920 году Ламб добавил к этому полное описание скелета NMC 2289 . Скелет NMC 2288 остаётся неописанным, и его кости по-прежнему хранятся в гипсе .

В 1920-е годы названия по канадским находкам получили ещё два вида, позднее классифицированные как эдмонтозавры, но в то время приписанные к роду Thespesius . Гилмор ввёл название Thespesius edmontoni в 1924 году, также основываясь на находке из формации каньона Хорсшу — почти полном скелете NMC 8399, у которого недоставало только большей части хвоста. NMC 8399 был обнаружен на реке Ред-Дир в 1912 году экспедицией Штернбергов . Ламб кратко описал его передние конечности, оссифицированные сухожилия и отпечатки кожи в 1913 и 1914 годах, первоначально посчитав его экземпляром вида, который он называл Trachodon marginatus , но затем изменив точку зрения . Этот скелет динозавра стал первым, выставленным на обозрение в канадских музеях. Гилмор определил, что новый вид близок к виду, который он называл Thespesius annectens , но различия в строении передних конечностей не позволили ему их объединить. Гилмор также отметил, что у T. edmontoni больше спинных и шейных позвонков, чем у экземпляра, описанного Маршем, но объяснил эти различия ошибкой Марша, по его мнению, переоценившего полноту доступного костного материала для этих частей тела Claosaurus annectens .
В 1926 году (ещё один сын Чарльза Штернберга-старшего) ввёл видовое название Thespesius saskatchewanensis для особи NMC 8509 — черепа и частичного скелета из окрестностей Рок-Крика на юге провинции Саскачеван . Этот материал был собран в 1921 году в отложениях, которые Штернберг классифицировал как Лансскую формацию (в настоящее время их относят к ). В костный материал NMC 8509 входили почти полный череп, большое количество позвонков, части плечевого и тазового поясов и частично сохранившиеся задние конечности; это был первый относительно хорошо сохранившийся динозавр, найденный в Саскачеване. Штернберг отнёс новый вид к роду Thespesius , поскольку в то время это был единственный род гадрозавридов, известный по ископаемым остаткам из Лансской формации , однако T. saskatchewanensis отличался необычно маленькими для этого рода размерами — ориентировочно от 7 до 7,3 м в длину .
От анатозавра до наших дней

В 1942 году, пытаясь упростить запутанную таксономию гадрозавров, Лалл и Райт ввели новое родовое название — Anatosaurus ; в этот род они включили виды, текущая родовая принадлежность которых была проблематичной. В качестве типового вида для анатозавров («уткоящеров», ot лат. anas , «утка», и др.-греч. σαῦρος , «ящерица»), получивших название за широкие, похожие на утиные клювы, был избран Claosaurus annectens Марша. В этот же род были включены:
- Thespesius edmontoni ,
- T. saskatchewanensis ,
- Trachodon longiceps , известный по большой нижней челюсти и получивший видовое название от Марша ещё в 1890 году,
- а также новый вид Anatosaurus copei , описанный по двум скелетам из Американского музея естественной истории, долгое время приписывавшимся к виду Diclonius mirabilis .
После изменения родового имени виды стали называться соответственно Anatosaurus annectens , A. edmontoni , A. saskatchewanensis , A. longiceps и A. copei . Анатозавров позже стали называть «классическими утконосыми динозаврами» .
Предложенная Лаллом и Райт классификация просуществовала несколько десятков лет с модификациями (так, в 1967 году Расселл и Чамни предложили объединить Anatosaurus edmontoni и Anatosaurus saskatchewanensis с E. regalis ), пока в 1970-е и 1980-е годы Майкл Бретт-Серман в рамках учёбы в университете не произвёл ревизию доступного материала. Он пришёл к выводу, что типовой вид анатозавров, A. annectens , на самом деле является эдмонтозавром и что A. copei отличается от других видов настолько, что должен быть выделен в отдельный род . В диссертации 1989 года Бретт-Серман предложил новое родовое имя для A. copei — Anatotitan , и в 1990 году зарегистрировал его вместе с Ральфом Чапманом . Два из трёх оставшихся видов, Anatosaurus saskatchewanensis и Anatosaurus edmontoni , также были причислены к эдмонтозаврам , а Anatosaurus longiceps — к анатотитанам, либо как отдельный вид Anatotitan longiceps , либо как синоним Anatotitan copei . Поскольку типовой вид анатозавров стал считаться эдмонтозавром, имя Anatosaurus больше не используется, считаясь синонимичным родовому имени Edmontosaurus .

В конечном итоге сформировалась концепция рода Edmontosaurus , включавшего три валидных вида: типовой E. regalis , E. annectens (включая Anatosaurus edmontoni/edmontonensis ) и E. saskatchewanensis . Дискуссия о таксономической принадлежности Anatotitan copei ещё продолжается: вернувшись к выводам Хетчера, сделанным ещё в 1902 году, спустя век, Джек Хорнер, и Кэтрин Форстер в 2004 году снова предложили рассматривать скелеты A. copei как экземпляры E. annectens с расплющенными черепами .
В 2007 были опубликованы данные о новой мумии. Этот экземпляр обнаружил в 1999 году Тайлер Лайсон в формации Хелл-Крик в Северной Дакоте, и после публикации мумия получила прозвище «Дакота» . В 2011 году Николас Кампион и Дэвид Эванс провели первый в истории морфометрический анализ по сравнению известных экземпляров, причисляемых к роду Edmontosaurus . Согласно выводам Кампиона и Эванса, валидными являются только два из трёх видов — E. regalis , из верхнего кампанского яруса , и E. annectens , из верхнего маастрихтского яруса . По мнению Кампионе и Эванса, их результаты подтверждают, что Anatotitan copei синонимичен E. annectens , в частности, что длинный, низкий череп A. copei представляет собой не более чем онтогенетическую вариацию у взрослых экземпляров E. annectens .
Видовой состав и распространение

Согласно последним данным, известны два валидных вида эдмонтозавров: типовой вид Edmontosaurus regalis и E. annectens . E. regalis известен только по окаменелостям из формации каньона Хорсшу в Альберте (Канада), которая относится к верхнему кампанскому и нижнему маастрихтскому ярусам позднего мелового периода. Известны ископаемые остатки более десяти представителей этого вида, в том числе семь скелетов с черепами и от пяти до семи отдельных черепов . В настоящее время считается, что экземпляры, причислявшиеся ранее к виду Thespesius edmontoni (или Anatosaurus edmontoni ), являются неполовозрелыми особями E. regalis . Trachodon cavatus и Agathaumas milo считаются потенциальными синонимами этого видового имени .
E. annectens известен по остаткам из формации Френчман (Саскачеван), формации Хелл-Крик (Монтана) и Лансской формации (Южная Дакота и Вайоминг). Все известные экземпляры относятся к верхнему маастрихтскому ярусу и включают не менее 20 черепов, с несколькими из которых найдены и другие костные остатки. В 1994 году Крейг Дерстлер назвал E. annectens «вероятно, самым изученным динозавром к настоящему времени» Экземпляры, ранее классифицировавшиеся как Anatosaurus copei и E. saskatchewanensis , сейчас считаются представителями E. annectens на разных стадиях развития — взрослыми ( A. copei ) и неполовозрелыми ( E. saskatchewanensis ) . Trachodon longiceps также считается синонимом E. annectens ; иногда также ставится знак равенства между Anatosaurus edmontoni ( Thespesius edmontonensis ) и E. annectens , но эта точка зрения оспаривается авторами, связывающими Anatosaurus edmontoni с E. regalis .
E. annectens отличался от E. regalis более длинным и низким, но менее мощным черепом Хотя Бретт-Серман предполагал, что E. regalis и E. annectens могут быть соответственно самцами и самками одного и того же вида , это маловероятно, так как все ископаемые остатки E. regalis были найдены в более ранних геологических формациях, чем остатки E. annectens .
Существуют также окаменелости, относимые к роду эдмонтозавров, видовая принадлежность которых не определена. Такие остатки (обозначаемые Edmontosaurus sp.) позволяют сделать выводы о распространении рода от Аляски (где предполагаемые остатки E. saskatchewanensis обнаружены в нижнемаастрихтской ) до Техаса, где аналогичные находки сделаны в .
Широкая область распространения эдмонтозавров — от Аляски до Колорадо, включая полярные области, где в течение большой части года светлое время суток сведено к минимуму, — стала причиной гипотез об их регулярных миграциях . Обзор исследований миграций динозавров, подготовленный в 2008 году, включает вывод о том, что E. regalis мог совершать ежегодные переходы, общая протяжённость которых доходила до 2600 км при наличии соответствующего обмена веществ и жировых запасов. Такая дальность миграций, которые позволяли бы животным доходить от Аляски до Альберты, подразумевает среднюю скорость движения от 2 до 10 км/ч. Предполагаемый кочевой образ жизни E. regalis отличает его от других видов эдмонтозавров и многих других динозавров — теропод , зауропод и анкилозавров , которые, согласно выводам авторов обзора, скорее всего проводили лето и зиму в одних и тех же местах . Противоположный вывод делают авторы исследования 2012 года, согласно которому микроструктура костей эдмонтозавров из формации Принс-Крик (Аляска) свидетельствует о зимовке в полярных областях .
Палеоэкология
Род эдмонтозавров был распространён на большой территории в течение длительного времени. Окаменелости, по которым он известен, могут быть разделены на две группы — более древние, из формаций каньона Хорсшу и Сент-Мери-Ривер, и более молодые, из Лансской формации, а также формаций Френчман и Хелл-Крик. Период, к которому относятся формации, подобные формации Хорсшу, известен также как эдмонтонский, а период, охватывающий более поздние находки, — как лансский. Динозавровая фауна, характерная для эдмонтонского и лансского периодов, различается .
Эдмонтонская палеоэкология

Ископаемые остатки Edmontosaurus regalis впервые появляются среди окаменелостей наземных позвоночных эдмонтонского яруса . Хотя иногда формация каньона Хорсшу описывается как исключительно раннемаастрихтская , на самом деле она охватывает несколько бо́льший период времени. Самые ранние отложения формации датируются верхним кампанским ярусом (примерно 73 млн лет назад), а самые поздние — 68—67,6 млн лет назад . Остатки Edmontosaurus regalis встречаются в самом нижнем из пяти слоёв формации, но отсутствуют как минимум в двух верхних . Эдмонтозаврам может принадлежать до трёх четвертей всех известных остатков в бедленде близ Драмхеллера (Альберта) . Считается, что в формации каньона Хорсшу большую роль играют морские отложения — в описываемый период этот регион занимало Западное внутреннее море , мелкое эпиконтинентальное море , на протяжении большей части мелового периода располагавшееся в пределах Североамериканского континента . Эту среду обитания эдмонтозавры делили с такими гадрозавридами, как зауролофы и гипакрозавры ; гипсилофодонтами рода Parksosaurus ; рогатыми динозаврами — монтаноцератопсами , анхицератопсами , арриноцератопсами и пахиринозаврами ; пахицефалозаврами стегоцерасами ; анкилозавридами эвоплоцефалами ; нодозавридами эдмонтониями ; орнитомимами и струтиомимами ; малоизученными мелкими тероподами из семейств троодонтидов и дромеозавридов ; а также тираннозавридами — альбертозаврами и дасплетозаврами . Эдмонтозавры населяли прибрежные зоны, тогда как другие гадрозавриды этого региона держались в низменных местностях дальше от моря . Типичная среда обитания эдмонтозавров представляла собой болотистые местности, поросшие болотным кипарисом , и верховые болота в дельтах рек. Такие же условия предпочитали и пахиринозавры, тогда как пойменные луга дальше от морского побережья занимали гипакрозавры, зауролофы, анхицератопсы и арриноцератопсы ; поэтому остатки эдмонтозавров и зауролофов вместе обычно не встречаются , в то время как общие скопления эдмонтозавров и пахиринозавров известны по прибрежным отложениям эдмонтонского яруса вплоть до Аляски .
Лансская палеоэкология
Лансский период в истории животного мира Северной Америки, завершившийся примерно 65,5 миллионов лет назад , был последним перед мел-палеогеновым вымиранием , в ходе которого исчезли все динозавры. Эдмонтозавры были в этот период одними из самых распространённых динозавров региона. Роберт Баккер сообщает, что кости эдмонтозавров составляли 1/7 от общего числа находок в большой выборке (из остальных находок 5/6 составляли кости трицератопсов ) . Регион сосуществования трицератопсов и эдмонтозавров на прибрежных равнинах простирался от Колорадо (Денверская формация) до Саскачевана (формация Френчман) . Соседями E. annectens по ареалу были также гипсилофодонтиды тесцелозавры , более редкие рогатые динозавры торозавры , травоядные пахицефалозавры и анкилозавры , а также тероподы — орнитомимы , троодоны и тираннозавры .

Формация Хелл-Крик, к которой относятся обнажённые породы в районе водохранилища Форт-Пек в Монтане, как считается, представляла собой поросшую лесом плоскую низменность с относительно сухим субтропическим климатом, создававшим благоприятные условия для разнообразных растений, включавших покрытосеменные деревья, хвойные деревья (в частности таксодиум двурядный ), папоротники и гинкго . Эти места находились в сотнях километров к западу от береговой линии. В животном мире были широко представлены речные черепахи и ведшие древесный образ жизни многобугорчатые млекопитающие , а на суше — гигантские вараны, по размерам не уступавшие комодским драконам . Из крупных динозавров самыми распространёнными были трицератопсы, а из небольших травоядных динозавров — тесцелозавры. Здесь в песчаных речных руслах сохранились ископаемые остатки эдмонтозавров, включая метрового детёныша, вероятно, принадлежавшие особям, погибшим при перекочёвке своего стада .
Лансская формация, представленная выходами на поверхность в 100 км к северу от Форт-Ларами (Вайоминг) , предположительно представляла собой байу наподобие существующих в настоящее время на прибрежных равнинах Луизианы. Она была ближе к устью большой реки, чем расположенная северней формация Хелл-Крик, и более богата наносными отложениями. В покрывавших эту равнину смешанных лесах встречались и тропические деревья — араукарии и пальмы , что отличало их от лесов лежащей северней прибрежной равнины . Во влажном субтропическом климате на заболоченных землях процветали сабалевые пальмы , хвойные деревья и папоротники, а на более сухих почвах леса образовывали хвойные деревья, ясени , вечнозелёные дубы и кустарники . Бок о бок с динозаврами эти места населяли пресноводные рыбы, саламандры, черепахи, змеи, береговые птицы и мелкие млекопитающие. В отличие от формации Хелл-Крик, здесь не было такого множества мелких динозавров, но тесцелозавры, по-видимому, были распространены и в этом регионе, а от трицератопсов остались многочисленные черепа, несколько менее крупные, чем у северных родичей. Именно в Лансской формации были обнаружены две «мумии» эдмонтозавров .
Палеобиология
Процесс роста
В 2011 году Кампионе и Эванс использовали параметры всех известных черепов эдмонтозавров кампанского и маастрихтского ярусов для создания морфометрического графика, сравнивающего размеры и другие характеристики черепов. Результаты исследования показали, что в ряде случаев в прошлом новые виды описывались на основе характеристик, напрямую связанных с размером черепа. Это позволило Кампионе и Эвансу сделать вывод о том, что череп эдмонтозавра существенно удлинялся и уплощался по мере взросления, что и привело к ошибочным классификациям в прошлом. Так, кампанский экземпляр Thespesius edmontoni (название считалось синонимом E. annectens из-за сходства в форме и размере черепа), скорее всего является подростковой особью E. regalis , а три ранее описывавшихся вида маастрихтских эдмонтозавров, по-видимому, представляют собой этапы взросления одного вида: E. saskatchewanensis — детёнышей, E. annectens — неполовозрелых особей и Anatotian copei — взрослых динозавров .
Питание

Как и остальные гадрозавриды , эдмонтозавры были крупными сухопутными травоядными ящерами. Их постоянно сменяющиеся зубы были скомпонованы в батареи; каждая батарея содержала сотни зубов, которые частично накладывались на соседние. Лишь небольшая часть зубов использовалась в каждый конкретный момент, равномерно стачиваясь по всей поверхности . Широкий «клюв» использовался для срывания стеблей и побегов — возможно, они срезались, как ножницами, вторым «лезвием» которых служили зубы нижней челюсти , или же челюсти смыкались, как створки раковины у моллюска, и потом со стебля обрывались более нежные листья и побеги . Поскольку в зубных батареях по краям челюстей есть глубокие выемки, предполагается, что у эдмонтозавров, как и у других птицетазовых динозавров , были своеобразные «щёки» , возможно, включавшие мышечные ткани (для некоторых видов были даже реконструированы схемы расположения таких мышечных тканей). Задачей этих щёк было удерживать пищу во рту . Эдмонтозавры, как и другие гадрозавриды, по-видимому, питались растительностью на высотах от уровня почвы до 2 метров, но могли доставать и листья с высоты до четырёх метров .
Со времени описания Джозефом Лейди в 1858 году вида Hadrosaurus foulkii и вплоть до 1970-х годов среди палеонтологов господствовало представление о гадрозавридах как о полуводных динозаврах, питающихся водяными растениями . Примером такой трактовки может служить описание черепа эдмонтозавра с сохранившимися остатками некостного материала клюва, сделанное в 1970 году Уильямом Моррисом. На этом основании Моррис делает вывод, что эдмонтозавры, наподобие современных уток, отцеживали клювом водяные растения и мелких моллюсков и рачков, сбрасывая воду через Л-образные щели в его верхней части . Эта интерпретация была отвергнута, поскольку щели в клювах эдмонтозавров больше напоминают аналогичные образования в клювах растительноядных черепах, чем мягкие образования, характерные для птиц, питающихся отфильтрованными водорослями и планктоном .
С середины 1980-х годов и до первого десятилетия XXI века представления об образе питания гадрозавридов в основном базировались на модели, предложенной в 1984 году Дэвидом Вейсхемпелом. Согласно этой модели, устройство черепа гадрозавридов делало возможным взаимное движение костей, обеспечивающее движение нижней челюсти вперёд и назад и изгиб наружу костей, несущих зубы, когда рот был закрыт. При таком устройстве зубы верхней челюсти могут скрести по зубам нижней, как рашпиль , перетирая попавшие между ними растительные волокна . В целом описываемый процесс, хотя и основываясь на иной анатомии, по достигаемому результату соответствует жевательному процессу у млекопитающих . В первом десятилетии XXI века, однако, появились работы, ставившие под сомнение выводы Вейсхемпела. Так, в 2008 году Холлидей и Уитмер показали, что у эдмонтозавров отсутствовали суставы, обеспечивающие взаимное движение костей черепа (такие суставы существуют, например, у чешуйчатых и птиц). По предположению Холлидея и Уитмера, части черепа, которым Вейсхемпел приписал эту функцию, в действительности представляли собой всего лишь хрящи . Важной деталью в модели Вейсхемпела было направление царапин на поверхности зубов, указывающее направление движения челюстей, но похожие следы могут быть оставлены и движениями другого вида, в частности, взаимным движением костей, составляющих нижнюю челюсть. Не все возможные модели были рассмотрены с использованием современной исследовательской техники . В 2009 году была опубликована работа Уильямса и коллег, посвящённая характеру износа зубов гадрозавридов. В работе показаны четыре вида царапин на зубах эдмонтозавров; самые распространённые, как указывается в публикации, связаны с движением в плоскостях, не параллельных друг другу, а не с простым движением вверх и вниз или вперёд и назад, как в модели Вейсхемпела. Два других вида царапин соответствуют движениям челюсти вперёд и назад, описанным Вейсхемпелом, а четвёртый тип, видимо, связан с открыванием рта. В целом рисунок царапин соответствует более сложному механизму перетирания пищи, чем предполагалось до этого .


Модель Вейсхемпела была разработана на основе компьютерной симуляции. В дальнейшем эта модель была усовершенствована Наталией Рыбчински и коллегами с помощью трёхмерной анимации, построенной на лазерном сканировании черепа эдмонтозавра. В новой модели предложенная Вейсхемпелом модель движения также воспроизводится, но при наличии дополнительного движения между другими костями черепа, некоторые из которых при жевании должны были расходиться на расстояние в 1,3—1,4 см. Рыбчински и коллеги, сомневаясь в жизнеспособности модели Вейсхемпела, тем не менее внесли в неё некоторые усовершенствования, включив в расчёты мягкие ткани и следы износа на зубах, лучше показывающие направление движений. Они указывают, что необходима проверка альтернативных гипотез . В публикации 2012 года Робин Катбертсон и коллеги называют модель Вейсхемпела маловероятной, в качестве альтернативы разбирая модель, в которой перетирание пищи происходит при движении нижней челюсти. Соединение нижней и верхней челюсти позволяет движение вперёд и назад вместе с обычными отклонениями; также предполагается, что подвижным было переднее соединение половин нижней челюсти. В целом новая модель предполагает небольшие движения двух половин нижней челюсти вперёд и назад одновременно с небольшими поворотами вдоль длинной оси. Такие движения лучше соответствуют описанным следам износа зубов и дают возможность более прочного устройства черепа, чем модель Вейсхемпела .
Поскольку царапины являются основной деталью износа зубов эдмонтозавра, Уильямс и коллеги в 2009 году предположили, что основу питания эдмонтозавра составляли жёсткие стебли, а не трава и листья — в последнем случае благодаря более нежному материалу царапин было бы меньше. Предполагается, что основной износ зубов связан с пережёвыванием таких богатых кремнезёмом растений, как хвощи , а также с попаданием на зубы земли в силу того, что часть пищи подбиралась с поверхности . Структура зубов показывает, что они могли использоваться как для срезания, так и для перетирания растительного материала .
Ранее сообщалось об обнаружении в желудке гадрозаврида клаозавра гастролитов — камешков, помогающих переваривать пищу , но это сообщение, вероятно, было ошибочным. Прежде всего, описанный экземпляр в настоящее время рассматривают как особь E. annectens ; в 1900 году, когда он был описан Б. Брауном , E. annectens считался видом клаозавров. Сами же камни, вероятнее всего, представляют собой обычный гравий, занесённый в костные остатки течением уже после смерти животного .
Содержание желудка

В процессе исследования обеих «мумий», найденных семьёй Штернбергов, сообщалось о возможном содержимом желудков. Для экземпляра, хранящегося в Американском музее естественной истории, о карбонизированном содержимом желудка сообщал Чарльз Штернберг, но это содержимое так никогда и не было описано .
Растительные остатки в тканях мумии из Зенкенбергского музея были описаны, но их оказалось трудно интерпретировать. Они включали хвою кипариса , сучья хвойных и лиственных деревьев и множество мелких семян или плодов . После появления описания этих остатков оно обсуждалось в немецкоязычном журнале Paläontologische Zeitschrift . Описавший эти материалы Рихард Крейзель рассматривал их как содержимое желудка , однако Отенио Абель не исключал возможности, что они были нанесены водой в труп животного после его смерти . В 1920-е годы гадрозавры считались животными, ведшими водный образ жизни, и Крейзель специально подчёркивал, что находки не исключают вероятности того, что гадрозавриды питались водяными растениями . Его находка, однако, осталась почти незамеченной в англоязычном научном сообществе, за исключением одного краткого упоминания о водно-сухопутной дихотомии , пока в 1960-е годы о ней не вспомнил Джон Остром в статье, пересматривающей теорию о ведущих водный образ жизни гадрозавридах. Вместо того, чтобы, как Крейзель, пытаться соотнести находку с водным образом жизни, Остром использовал её как доказательство того, что гадрозавриды были сухопутными травоядными . Хотя палеонтологическое сообщество в дальнейшем склонилось к позиции Острома , происхождение растительных остатков из зенкенбергской мумии по-прежнему остаётся под вопросом. По мнению палеонтолога- тафономиста Кеннета Карпентера , они могут представлять собой содержимое желудка умирающего от голода животного, а не его обычную диету . Фастовский и Смит в 2004 году писали, что, ввиду преобладания покрытосеменных растений в конце мелового периода, наличие семян и хвои голосеменных растений в желудке может свидетельствовать о том, что эдмонтозавр предпочитал именно такую пищу . Другие авторы отмечали, что, поскольку растительные остатки были извлечены из тела и препарированы , подтвердить выводы из первой публикации Крейзеля невозможно, и вероятность того, что они были занесены в труп после смерти животного, не может быть исключена .
Изотопный анализ зубов
Диета и физиология эдмонтозавров также оценивались по устойчивым изотопам углерода и кислорода в зубной эмали . Кислород и углерод, поступающие в организм с дыханием, едой и питьём, остаются в костях животных. Изотопы этих двух элементов определяются множеством внешних и внутренних факторов, таких, как виды употребляемых в пищу растений, физиология животного, солёность воды и климат. Если соотношение изотопов в окаменелостях не подвергалось изменениям в результате фоссилизации и диагенеза , оно может быть использовано для получения информации о факторах, его сформировавших. Для этих целей чаще всего используется зубная эмаль, поскольку структура формирующего её минерала превращает её в более устойчивую к последующим химическим изменениям ткань, чем кость и дентин .
В исследовании 2004 года К. Томас и С. Карлсон использовали зубы верхней челюсти от трёх особей, классифицированных как детская, подростковая и взрослая. Эти ископаемые остатки были найдены в костеносном слое формации Хелл-Крик в округе Корсон (Южная Дакота). В процессе исследования костный материал извлекался из следующих один за другим зубов в зубных батареях каждой особи с помощью микросвёрл. Этот метод использует специфику строения зубных батарей гадрозавридов для того, чтобы проследить изменения в соотношении изотопов с течением времени. Исследование показывает, что новые зубы у эдмонтозавра формировались чуть меньше чем за 2/3 года (несколько быстрей у более молодых особей). Зубы всех трёх экземпляров демонстрируют изменения в пропорции изотопов кислорода, возможно, соответствующие смене тёплых и сухих периодов более холодными и влажными; Томас и Карлсон также рассматривали возможность, что эти изменения вызваны миграциями, но оценивают связь со сменой времён года как более вероятную: миграции, по их мнению, скорей привели бы к унификации пропорций, поскольку большинство животных мигрирует так, чтобы оставаться в определённом температурном режиме или вблизи от излюбленных источников пищи .
Для зубов эдмонтозавров оказалось также характерным сочетание изотопов углерода, которое у млекопитающих указывало бы на смешанную диету из растений с разными видами метаболического связывания углерода — трёхатомным, как у большинства растений, и четырёхатомным, как у травянистых растений. Поскольку трава, для которой типично четырёхатомное связывание углерода, была редкостью в верхнем меловом периоде (если вообще существовала), Томас и Карлсон предлагают несколько альтернативных факторов, которые могли сформировать такое соотношение изотопов. По их мнению, наиболее вероятными факторами являются повышенное содержание в диете голосеменных ; употребление в пищу содержащих избыточное количество солей растений из прибрежных районов Западного внутреннего моря ; или физиологические отличия динозавров от млекопитающих, из-за которых соотношение углеродных изотопов в скелете динозавров шло иначе, чем у млекопитающих. Возможно также сочетание нескольких из этих факторов .
Патологии
В 2003 году была опубликована информация об опухолях на костной ткани эдмонтозавра, включавших гемангиомы , десмопластическую фиброму , раковые метастазы и . Эти результаты стали доступны после того, как позвонки динозавров были подвергнуты в поисках опухолей компьютерной томографии и флюороскопированию . Опухоли также были обнаружены у трёх других гадрозавридов — брахилофозавра , гилмореозавра и бактрозавра . Хотя были проанализированы более десяти тысяч окаменелостей, опухоли были найдены только у эдмонтозавров и близких к ним родов. Причиной опухолей, таким образом, могли быть как внешние факторы, так и генетическая предрасположенность (хотя отсутствие опухолей у других динозавров может быть связано просто с недостаточным объёмом материала — у эдмонтозавров такие опухоли встречаются в среднем только на одном из каждых 500 позвонков) .
У эдмонтозавров также был обнаружен остеохондроз — углубления на внешней поверхности костей в местах их соединения. Эта характеристика, связанная с тем, что хрящевой материал при росте не заменяется костным при росте, найдена в 2,2 % из 224 исследованных фаланг пальцев эдмонтозавров. Причины заболевания остаются неизвестными и могут быть связаны с генетической предрасположенностью, травмами, особенностями рациона, перепадами в кровоснабжении, излишком тиреоидных гормонов или нехваткой других гормонов, влияющих на рост организма. Из всех динозавров остеохондроз, как и опухоли, встречается чаще всего у гадрозавридов .
Способ передвижения

Считается, что для эдмонтозавров, как и для других гадрозавридов, был характерен факультативный бипедализм : в основном они передвигались на четырёх ногах, но при необходимости могли подниматься на задние. Вероятно, эдмонтозавры опирались на четыре ноги, когда стояли или передвигались медленно, а на задние ноги поднимались, когда двигались быстрее . Компьютерные модели 2007 года показывают, что эдмонтозавр мог развивать скорость до 45 км/ч ; дальнейшее моделирование для подростковой особи, при жизни весившей около 715 кг, показывает возможность передвигаться рысью , иноходью , шагом или галопом . К удивлению исследователей, модели показали, что наиболее быстрым способом передвижения были кенгуроподобные скачки́, позволявшие развивать скорость до 17,3 м/с. Эту модель передвижения, однако, учитывая габариты животного и отсутствие окаменелых следов, соответствующих скачка́м на двух ногах, исследователи считают маловероятной и демонстрирующей несовершенство компьютерной симуляции. Не считая скачков на двух ногах, самыми быстрыми способами передвижения, согласно моделям, были галоп (максимальная скорость в симуляции 15,7 м/с) и бег на задних ногах (до 14 м/с). Имеющиеся данные поддерживают версию о беге на задних ногах как наиболее вероятном способе быстрого передвижения, но авторы не исключают и возможности, что и в этих случаях эдмонтозавры могли двигаться на всех четырёх ногах .
Хотя гадрозавриды долгое время считались животными, ведшими водный или полуводный образ жизни, тело эдмонтозавров было хуже приспособлено для плавания, чем тела других динозавров, в частности, теропод , которые, как раньше считалось, были неспособны преследовать их в водной среде. Для гадрозавридов характерны относительно тонкие передние конечности с короткими пальцами, что делает их малоподходящими для плавания, а хвост не мог эффективно выполнять функцию движителя в воде, поскольку оссифицированные сухожилия лишали его необходимой гибкости. Слаборазвитые узлы крепления хвостовых мышц также показывают, что эдмонтозавры не могли эффективно размахивать хвостом из стороны в сторону .
Сосуществование с тероподами

Период существования и ареал эдмонтозавров пересекаются с эпохой существования и ареалом тираннозавров . При этом, как отмечает Джон Остром, у этого вида отсутствуют явно выраженные механизмы защиты от хищников: эдмонтозавры не были вооружены эффективными в бою шипами, когтями или зубами, но при этом не были и достаточно быстрыми, чтобы убежать от хищника (более поздние компьютерные модели показывают, что эдмонтозавры могли бегать как минимум быстрее тираннозавров ). Остром выдвигает предположение, что хорошо развитое обоняние предупреждало эдмонтозавров о приближающейся опасности задолго до столкновения, позволяя им загодя отступить в воду, где преследование становилось затруднительным. Остром, критиковавший теории о водном образе жизни эдмонтозавров, таким образом объясняет наличие перепонок между пальцами на передних конечностях «мумии траходона» .
Хвостовая часть скелета E. annectens , демонстрируемого в Денверском музее природы и науки, носит следы укуса теропода : 13-й и 14-й позвонки, считая от бедра, имеют повреждения, показывающие, что нападение совершалось справа и сзади. Часть остистого отростка одного из позвонков срезана, а другие загнуты, причём на трёх присутствуют следы зубов. Поскольку основание хвоста располагалось на высоте не менее 2,9 метра, единственным тероподом из этой геологической формации, способным нанести такие травмы, был Tyrannosaurus rex . Позвонки частично зажили, но их обладатель умер прежде, чем следы укуса исчезли окончательно; кости также носят следы инфекции. Изучавший данный экземпляр Кеннет Карпентер отмечает, что левое бедро носит следы перелома, более раннего, чем укусы в районе хвоста, поскольку процесс его заживления зашёл дальше. Это позволило Карпентеру предположить, что охромевший из-за более ранней травмы эдмонтозавр стал удобной мишенью для атаки. С другой стороны, поскольку эдмонтозавр выжил в этом нападении, Карпентер предполагает, что ему удалось сбежать от хищника или даже что повреждения хвоста были вызваны тем, что гадрозаврид использовал его как оружие против тираннозавра, размахивая им из стороны в сторону .
Нижняя челюсть другого экземпляра E. annectens из Южной Дакоты длиной 7,6 метра носит следы зубов мелких теропод. Некоторые из ран частично зажили. Майкл Триболд, описывавший данный образец, предположил, что мелкие тероподы пытались добраться до горла эдмонтозавра, травоядный ящер выжил в этом нападении, но умер вскоре после этого от полученных ран .
Некоторые из известных кладбищ эдмонтозавров посещались падальщиками. Следы зубов альбертозавров и заурорнитолестов характерны для одного из костеносных слоёв в Альберте , а другой сохранил свидетельства того, что трупами эдмонтозавров и зауролофов питался дасплетозавр .
Социальное поведение
Существование костеносных слоёв с большим количеством ископаемых остатков эдмонтозавров, как и других гадрозавридов, позволяет предполагать, что они были стадными животными . В базе данных 2007 года, содержащей информацию об ископаемых костеносных слоях, содержатся данные четырёх карьеров с остатками эдмонтозавров — на Аляске (формация Принс-Крик), в Альберте (формация каньона Хорсшу), в Южной Дакоте (хелл-крикская формация) и в Вайоминге (лансская формация) . Один из костеносных слоёв, в глинистых сланцах Лансской формации в восточном Вайоминге, занимает площадь более квадратного километра, хотя кости собственно эдмонтозавров встречаются в основном на участке площадью порядка 40 гектаров. Количество эдмонтозавров, которым принадлежат кости в этом месторождении, оценивается в 10—25 тысяч экземпляров . В середине XX века было отмечено, что и остатки молодых гадрозавридов, предположительно обитавших на прибрежных равнинах, в основном обнаруживаются в местах, соответствующих более возвышенным местностям. Это породило гипотезу о том, что гадрозавриды изначально появились на возвышенностях и в дальнейшем возвращались в родные края, чтобы откладывать яйца. Однако в дальнейшем отсутствие яиц и костей молодняка в низменных районах получило другое объяснение: кислотность почв рядом с морем была слишком высокой, и тонкие кости и скорлупа в них попросту не сохранялись .
У эдмонтозавров отсутствовал костяной гребень на голове, по-видимому, служивший другим гадрозавридам для усиления голоса, но вполне вероятно, что его роль выполняли конструкции из мягких тканей. Как отмечает Джеймс Хопсон, на костях черепа эдмонтозавров вокруг ноздрей заметны глубокие выемки, где, как предполагается, размещались надувающиеся мешки. Эти мешки, как считает Хопсон, служили для передачи как визуальных, так и, в особенности, звуковых сигналов, которые в условиях более лесистой местности, чем та, в которой обитали другие утконосые динозавры, могли иметь повышенное значение для поддержания контакта . Социальную роль, возможно, исполнял и гребень из мягких тканей на макушке, обнаруженный у E. regalis .
Возможно, что для эдмонтозавров был характерен диморфизм , при котором часть особей была сложена более мощно, чем остальные, но не установлено достоверно, был ли этот диморфизм половым .
Примечания
- ↑ (англ.) информация на сайте Paleobiology Database . (Дата обращения: 1 февраля 2022) .
- . Динозавры. Полный определитель = The Princeton Field Guide to Dinosaurs : ориг. изд. 2016 / пер. с англ. К. Н. Рыбакова ; науч. ред. А. О. Аверьянов . — М. : Фитон XXI, 2022. — С. 331—333 . — 360 с. : ил. — ISBN 978-5-6047197-0-1 .
- ↑ Sandeep Ravindran. . National Geographic (12 декабря 2013). Дата обращения: 22 апреля 2014. 25 апреля 2014 года.
- ↑ , p. 392.
- Matthew T. Carrano. // Amniote paleobiology: perspectives on the evolution of mammals, birds, and reptiles / Matthew T. Carrano, Timothy J. Gaudin, Richard W. Blob, John R. Wible (Eds.). — The University of Chicago Press, 2006. — P. 267. — ISBN 0-226-09477-4 .
- Horner, J. R., and Woodward, H. N. : [ 4 марта 2016 ] // Hadrosaur Symposium Abstracts. — Drumheller, AB : Royal Tyrrell Museum of Paleontology, 2011. — Vol. 1. — P. 66.
- ↑ , p. 94.
- , p. 560.
- ↑ , p. 225.
- ↑ , p. 440.
- ↑ Campione, N. E., Evans, D. C. (англ.) // PLOS One . — Public Library of Science , 2011. — Vol. 6, no. 9: e25186 . — doi : . 3 апреля 2012 года.
- Frederic A. Lucas. // Smithsonian Miscellaneous Collections. — 1904. — Vol. 45. — P. 317—320.
- ↑ William J. Morris. // Contributions in Science (Los Angeles County Museum of Natural History). — 1970. — Vol. 193. — P. 1—14.
- ↑ , p. 389.
- E. D. Cope. On the characters of the skull in the Hadrosauridae // Proceedings of the Philadelphia Academy of Natural Sciences. — 1883. — Vol. 35. — P. 97—107.
- , p. 5.
- , p. 152.
- ↑ , p. 443.
- ↑ , p. 113.
- ↑ , p. 394.
- ↑ Kathryn J. Stanton Thomas, Sandra J. Carlson. // Palaeogeography, Palaeoclimatology, and Palaeoecology. — 2004. — Vol. 206. — P. 257—287. — doi : . 28 марта 2014 года.
- , p. 68.
- , pp. 153, 156, 160.
- ↑ Erickson, Gregory M.; Krick, Brandon A.; Hamilton, Matthew; Bourne, Gerald R.; Norell, Mark A.; Lilleodden, Erica; and Sawyer, W. Gregory. (англ.) // Science. — 2012. — Vol. 338, no. 6103 . — P. 98—101. — doi : . 9 октября 2012 года.
- ↑ , p. 452.
- ↑ James A. Hopson. The evolution of cranial display structures in hadrosaurian dinosaurs (англ.) // . — , 1975. — Vol. 1. — P. 21—43. — ISSN .
- , pp. 128.
- , p. 153.
- , p. 162.
- , p. 453.
- ↑ , p. 462.
- Peter M. Galton. // Journal of Paleontology. — 1970. — Vol. 44, № 3 . — P. 464—473.
- ↑ John H. Ostrom . // American Journal of Science. — 1964. — Vol. 262, № 8 . — P. 975—997. — doi : .
- , pp. 87—90.
- , p. 109.
- , pp. 92—93.
- , pp. 82—83.
- , pp. 84—86.
- ↑ Henry Fairfield Osborn. (англ.) // Science. — 1909. — Vol. 29, no. 750 . — P. 793—795. — doi : .
- ↑ Henry Fairfield Osborn. // Memoirs of the American Museum of Natural History. — 1912. — Vol. 1. — P. 33—54. 21 декабря 2014 года.
- ↑ . National Geographic News (3 декабря 2007). Дата обращения: 5 августа 2013. Архивировано из 31 октября 2012 года.
- ↑ Christopher Lee. . The Washington Post (3 декабря 2007). Дата обращения: 5 сентября 2013. Архивировано из 13 октября 2008 года.
- ↑ Phillip L. Manning, et al. Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA) // Proceedings of the Royal Society B. — 2009. — Vol. 276. — P. 3429—3437. — doi : .
- . Московский Комсомолец (8 июня 2010). Дата обращения: 26 декабря 2012. Архивировано из 3 марта 2014 года.
- P. R. Bell, F. Fanti, P. J. Currie, and V. M. Arbour. A mummified duck-billed dinosaur with a soft-tissue cock's comb // Current Biology. — 2013. — Vol. 24, № 1 . — P. 70—75. — doi : .
- , pp. 112—114.
- ↑ Kenneth Carpenter. Paleoecological significance of droughts during the Late Cretaceous of the Western Interior // Fourth Symposium on Mesozoic Terrestrial Ecosystems, Drumheller, August 10–14, 1987 / Currie, Philip J. and Koster, Emlyn H. (Eds.). — Drumheller, AB: Royal Tyrrell Museum of Palaeontology, 1987. — Vol. 3. — P. 42—47. — (Occasional Paper of the Tyrrell Museum of Palaeontology). — ISBN 0-7732-0047-9 .
- Kenneth Carpenter. // The Journal of Paleontological Sciences. — 2007. — Vol. 1:JPS.C.07.0002. 23 августа 2009 года.
- , p. 111.
- ↑ Bakker, Robert T. The case of the duckbill's hand // The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. — NY, 1986. — P. 146—159. — ISBN 0-8217-2859-8 .
- , p. 114.
- John R. Horner . A "segmented" epidermal frill in a species of hadrosaurian dinosaur // Journal of Paleontology. — Vol. 58. — P. 270—271.
- Lyson, Tyler R., Hanks, H. Douglas, and Tremain, Emily S. // Abstracts with Programs — Geological Society of America. — 2003. — Vol. 35. 24 февраля 2016 года.
- O. C. Marsh . // American Journal of Science. — 1893. — Vol. 45. — P. 83—86. 15 ноября 2017 года.
- Othniel Charles Marsh . The dinosaurs of North America // . — Washington, D.C.: U.S. Geological Survey, 1896. — P. 133—244. 9 мая 2009 года. (осторожно, массивный PDF-файл — искать на с. 222)
- , pp. 51—56.
- Barnum Brown . Anchiceratops , a new genus of horned dinosaurs from the Edmonton Cretaceous of Alberta, with discussion of the ceratopsian crest and the brain casts of Anchiceratops and Trachodon (англ.) // . — American Museum of Natural History , 1914. — Vol. 33. — P. 539—548.
- , pp. 122—128.
- ↑ Jerison, Harry J., Horner, John R., and Horner, Celeste C. Dinosaur forebrains (англ.) // Journal of Vertebrate Paleontology . — . — Vol. 21, no. 3, Suppl. .
- ↑ Harry J. Jerison. // Encyclopedia of Neuroscience / Adelman, G. & Smith, B.H. (Eds.). — 3rd Edition. — Elsevier, 2004. — ISBN 0444514325 .
- , p. 51.
- ↑ , p. 127.
- , pp. 126.
- , pp. 127—128.
- ↑ Xing H., Mallon J. C., and Currie M. L. (англ.) // PLOS One . — Public Library of Science , 2017. — Vol. 12, no. 4 . — P. e0175253. — doi : . 17 июля 2017 года.
- ↑ Prieto-Márquez A. : [ 9 октября 2016 ] // Zoological Journal of the Linnean Society. — 2010. — Vol. 159. — P. 435—502. — doi : .
- ↑ , p. 459.
- Mori H., Druckenmiller P. S., and Erickson G. M. // Acta Palaeontologica Polonica . — Polish Academy of Sciences , 2016. — Vol. 61, № 1 . — P. 15—32. — doi : . 2 декабря 2017 года.
- Takasaki R., Fiorillo A. R., Tykoski R. S., and Kobayashi Y. (англ.) // PLOS One. — 2020. — Vol. 15, no. 5 . — P. e0232410. — doi : . 7 мая 2020 года.
- , p. 48.
- , p. 554.
- Gates T. A., and Sampson S. D. (англ.) // . — Oxford University Press , 2007. — Vol. 151, no. 2 . — P. 351—376. — doi : . 24 мая 2014 года.
- ↑ Creisler, Benjamin S. Deciphering duckbills: a history in nomenclature // Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs / Carpenter, Kenneth (ed.). — Bloomington and Indianapolis: Indiana University Press, 2007. — P. 185—210. — ISBN 0-253-34817-X .
- E. D. Cope. // Proceedings of the American Philosophical Society. — 1871. — Vol. 12, № 86 . — P. 41—52.
- ↑ , p. 557.
- E. D. Cope. // Bulletin of the United States Geological and Geographical Survey of the Territories. — 1874. — Vol. 1. — P. 9—28.
- ↑ Kenneth Carpenter and D. Bruce Young. // Rocky Mountain Geology. — 2002. — Vol. 37, № 2 . — P. 237—254. — doi : .
- E. D. Cope. // U.S. Geological and Geographical Survey of the Territories Annual Report. — 1874. — Vol. 2. — P. 429—454.
- Lambe, Lawrence M. On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous) // Contributions to Canadian Paleontology. — 1902. — Vol. 3. — P. 25—81.
- , p. 28.
- , p. 558.
- O. C. Marsh . // American Journal of Science. — 1892. — Vol. 43. — P. 449—453. 30 сентября 2017 года.
- O. C. Marsh . // American Journal of Science. — 1892. — Vol. 44, № 262 . — P. 343—349.
- C. E. Beecher. The Reconstruction of a Cretaceous Dinosaur, Claosaurus annectens Marsh // Transactions of the Connecticut Academy of Arts and Sciences. — 1902. — Vol. 11. — P. 311—323.
- Charles W. Gilmore. // Proceedings of the United States National Museum. — 1941. — Vol. 90. — P. 305—377. 27 апреля 2014 года.
- J. B. Hatcher. // Annals of the Carnegie Museum. — 1902. — Vol. 14, № 1 . — P. 377—386.
- Charles W. Gilmore. // Science . — 1915. — Vol. 41, № 1061 . — P. 658—660. — doi : . — . 18 июля 2018 года.
- ↑ Mark A. Norell, Eugene S. Gaffney, and Lowell Dingus. Edmontosaurus annectens // . — New York: Nevraumont Publishing, 1995. — P. —155. — ISBN 0-679-43386-4 .
- Cristiano Dal Sasso. . — Indiana University Press, 2004. — P. 112. — ISBN 0-253-34514-6 .
- ↑ Lawrence M. Lambe. // The Ottawa Naturalist. — 1917. — Vol. 31, № 7 . — P. 65—73.
- , p. 1.
- , pp. 1—5.
- , p. 390.
- , p. 161.
- Lawrence M. Lambe. // The Ottawa Naturalist. — 1913. — Vol. 27. — P. 21—25.
- Lawrence M. Lambe. // The Ottawa Naturalist. — 1914. — Vol. 27. — P. 129—135.
- Charles W. Gilmore. A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta. — Department of Mines, Geological Survey of Canada, 1924. — Vol. 38. — P. 13—26. — (Bulletin).
- , pp. 159—160.
- Charles M. Sternberg. A new species of Thespesius from the Lance Formation of Saskatchewan. — Department of Mines, Geological Survey of Canada, 1926. — Vol. 44. — P. 77—84. — (Bulletin).
- , p. 160.
- , pp. 151—164.
- Donald F. Glut. . — Secaucus, NJ: Citadel Press, 1982. — ISBN 0-8065-0782-9 .
- M. K. Brett-Surman. (англ.) // Nature . — 1979. — Vol. 277, no. 5697 . — P. 560—562. — doi : . 19 июля 2018 года.
- ↑ , p. 76.
- Ralph E. Chapman and Michael K. Brett-Surman. // Dinosaur Systematics: Perspectives and Approaches / Carpenter, Kenneth, and Currie, Philip J. (eds.). — Cambridge: Cambridge University Press, 1990. — P. 163—177. — ISBN 0-521-43810-1 .
- George Olshevsky. A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. — San Diego: Publications Requiring Research, 1991. — P. 170—171. — (Mesozoic Meanderings No. 2).
- , pp. 440, 460.
- ↑ Nicolas E. Campione, and David Evans. (англ.) // PLOS One . — Public Library of Science . — Vol. 6, no. 9: e25186 . — doi : . 3 апреля 2012 года.
- , p. 77.
- ↑ Behrensmeyer, Anna K. (Excel spreadsheet). Bonebeds: Genesis, Analysis, and Paleobiological Significance . University of Chicago Press (2007). Дата обращения: 7 декабря 2008. 30 июня 2019 года.
- , p. 582.
- P. R. Bell and E. Snively. . — 2008. — Vol. 32. — P. 271—284. — doi : . 19 апреля 2014 года.
- Robin Lloyd. . LiveScience.com (4 декабря 2008). Дата обращения: 30 марта 2014.
- A. Chinsamy, D. B. Thomas, A. R. Tumarkin-Deratzian, and A. R. Fiorillo. . — 2012. — Vol. 295, № 4 . — P. 610—614. — doi : . 26 мая 2014 года.
- Peter Dodson. . — Princeton, NJ: Princeton University Press, 1996. — P. 14. — ISBN 0-691-05900-4 .
- Robert M. Sullivan and Spencer G. Lucas. // Late Cretaceous Vertebrates from the Western Interior / Lucas, Spencer G.; and Sullivan, Robert M. (eds.). — Albuquerque, NM: New Mexico Museum of Natural History and Science, 2006. — P. 7—29. — (New Mexico Museum of Natural History and Science Bulletin No. 35).
- , pp. 440, 577.
- Xiao-chun Wu, Donald B. Brinkman, David A. Eberth, and Dennis B. Braman. // Canadian Journal of Earth Science. — 2007. — Vol. 44, № 9 . — P. 1243—1265. — doi : . 4 марта 2014 года.
- David A. Eberth. (2002). . 75th Anniversary of the CSPG Convention. Vol. 117. из оригинала 25 апреля 2014 . Дата обращения: 24 апреля 2014 .
- ↑ , pp. 170—174.
- D. G. Smith. // Geological Atlas of the Western Canada Sedimentary Basin / Mossop, G.D. and Shetsen, I. (comp.). — Canadian Society of Petroleum Geologists and Alberta Research Council, 1994.
- , p. 577.
- , p. 559.
- ↑ . ScienceDaily (3 июля 2007). Дата обращения: 28 февраля 2014. 9 ноября 2020 года.
- ↑ Thomas M. Lehman. Late Cretaceous dinosaur provinciality // / Tanke, Darren, and Carpenter, Kenneth (eds.). — Indiana University Press, 2001. — P. —328. — ISBN 0-253-33907-3 .
- (англ.) . Encyclopædia Britannica . Дата обращения: 27 апреля 2014.
- Bakker, Robert T. The twilight of the dinosaurs // The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. — NY, 1986. — P. 438. — ISBN 0-8217-2859-8 .
- , p. 578.
- Phillip Bigelow. (21 июля 2010). Дата обращения: 6 февраля 2014. 24 января 2007 года.
- , pp. 175—180.
- ↑ , pp. 180—181.
- Kraig Derstler. Dinosaurs of the Lance Formation in eastern Wyoming // The Dinosaurs of Wyoming / Nelson, Gerald E. (Ed.). — Wyoming Geological Association, 1994. — P. 127—146.
- ↑ David E. Fastovsky, and Joshua B. Smith. // / David B. Weishampel, Peter Dodson, Halszka Osmólska (Eds.). — 2nd ed. — Berkeley, CA: University of California Press, 2004. — P. —626. — ISBN 0-520-24209-2 .
- Peter M. Galton. The cheeks of ornithischian dinosaurs // Lethaia. — 1973. — Vol. 6. — P. 67—89. — doi : .
- ↑ Paul M. Barrett. // Palaeontology. — 2005. — Vol. 48, № 2 . — P. 347—358. — doi : . 7 декабря 2013 года.
- ↑ N. Rybczynski, A. Tirabasso, P. Bloskie, R. Cuthbertson, and C. Holliday. (англ.) // . — Coquina Press, 2008. — Vol. 11, no. 2 . 21 апреля 2012 года.
- ↑ Casey M. Holliday and Lawrence M. Witmer. (англ.) // Journal of Vertebrate Paleontology . — , 2008. — Vol. 28, no. 4 . — P. 1073—1088. — doi : . 18 июля 2018 года.
- ↑ V. S. Williams, P. M. Barrett, and M. A. Purnell. (англ.) // Proceedings of the National Academy of Sciences . — National Academy of Sciences , 2009. — Vol. 106, no. 27 . — P. 11194—11199. — doi : . 24 сентября 2015 года.
- R. S. Cuthbertson, A. Tirabasso, N. Rybczynski, & R. B. Holmes. // The Anatomical Record. — 2012. — Vol. 295, № 6 . — P. 968—979. — doi : . 22 мая 2014 года.
- Barnum Brown. Gastroliths (англ.) // Science. — 1907. — Vol. 25, no. 636 . — P. 392.
- ↑ J. S. Tweet, K. Chin, D. R. Braman, and N. L. Murphy. Probable gut contents within a specimen of Brachylophosaurus canadensis (Dinosauria: Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana (англ.) // . — , 2008. — Vol. 23, no. 9 . — P. 624—635. — doi : .
- Catherine Forster. Hadrosauridae // / Philip J. Currie, Kevin Padian (Eds.). — Academic Press, 1997. — P. 293—300. — ISBN 9780080494746 .
- Karen Chin. What did dinosaurs eat: Coprolites and other direct evidence of dinosaur diets // The complete dinosaur / M. K. Brett-Surman, Thomas R. Holtz Jr., and James O. Farlow (Eds.). — 2nd ed. — Bloomington, IN: Indiana University Press, 2012. — P. 589—602. — ISBN 978-0-253-00849-7 .
- Wieland, G. R. Dinosaur feed (англ.) // Science. — 1925. — Vol. 61, no. 1589 . — P. 601—603. — doi : . — .
- Michael J. Ryan and Matthew K. Vickaryous. Diet // Encyclopedia of Dinosaurs / P. J. Currie, K. Padiam (Eds.). — Acaedemic Press, 1997. — P. 169—174. — ISBN 978-0-12-116810-6 .
- B. M. Rothschild, D. H. Tanke, M. Helbling, and L. D. Martin. // Naturwissenschaften. — 2003. — Vol. 90, № 11 . — P. 495—500. — doi : . — .
- Bruce Rothschild and Darren H. Tanke. // Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs / Carpenter, Kenneth (ed.). — Bloomington and Indianapolis: Indiana University Press, 2007. — P. 171—183. — ISBN 0-253-34817-X .
- W. I. Sellers, P. L. Manning, T. Lyson, K. Stevens, and L. Margetts. (англ.) // . — Coquina Press, 2009. — Vol. 12, no. 3 . 9 марта 2014 года.
- , pp. 342—343.
- M.K Brett-Surman. Ornithopods // The Complete Dinosaur / James O. Farlow and M. K. Brett-Surman (eds.). — Bloomington: Indiana University Press, 1997. — P. 342—343. — ISBN 0-253-33349-0 .
- Kenneth Carpenter. // Gaia. — 1998. — Vol. 15. — P. 135—144. — ISSN . 26 сентября 2013 года.
- Campagna, Tony. The PT interview: Michael Triebold (неопр.) // Prehistoric Times. — 2000. — Т. 40 . — С. 18—19 .
- , p. 396.
- Aase Roland Jacobsen and Michael J. Ryan. Taphonomic aspects of theropod tooth-marked bones from an Edmontosaurus bone bed (Lower Maastrichtian), Alberta, Canada (англ.) // Journal of Vertebrate Paleontology . — , 1999. — Vol. 19, no. 3 suppl. . — P. 55A. — doi : .
- , p. 463.
- A. Chadwick, L. Spencer, and L. Turner. // Poster presented at SVP 66th Annual Meeting. — 2006. 5 марта 2014 года.
- , pp. 461—462.
- Rebecca Gould, Robb Larson, and Ron Nellermoe. An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD (англ.) // Journal of Vertebrate Paleontology . — , 2003. — Vol. 23, no. 3 suppl. . — P. 56A—57A.
Литература
- Michael Keith Brett-Surman. . — Washington, D.C.: George Washington University , 1989. — (Ph.D. dissertation).
- Richard J. Butler and Paul M. Barrett. Ornithopods // The complete dinosaur / M. K. Brett-Surman, Thomas R. Holtz Jr., and James O. Farlow (Eds.). — 2nd ed. — Bloomington, IN: Indiana University Press , 2012. — P. 551—568. — ISBN 978-0-253-00849-7 .
- Donald F. Glut. Edmontosaurus // . — Jefferson, NC: McFarland & Company , 1997. — P. —396. — ISBN 0-89950-917-7 .
- John R. Horner, David B. Weishampel and Catherine A. Forster. Hadrosauridae // / David B. Weishampel, Peter Dodson, Halszka Osmólska (Eds.). — 2nd ed. — Berkeley, CA: University of California Press , 2004. — P. —463. — ISBN 0-520-24209-2 .
- Lawrence Lambe. The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta. — Department of Mines, Geological Survey of Canada, 1920. — Vol. 120. — P. 1—79. — (Memoir). — ISBN 0-659-96553-4 .
- Richard Swann Lull, Nelda E. Wright. . — Geological Society of America , 1942. — (Geological Society of America Special Paper 40 ).
- Dale A. Russell. The 'Modern' Cretaceous // An Odyssey in Time: Dinosaurs of North America. — Minocqua, WI: NorthWord Press, 1989. — P. 170—193. — ISBN 1-55971-038-1 .
- David B. Weishampel and John R. Horner. Hadrosauridae // The Dinosauria / David B. Weishampel, Peter Dodson, Halszka Osmólska (Eds.). — Berkeley, CA: University of California Press, 1990. — P. 534—561. — ISBN 0-520-06726-6 .
- David B. Weishampel, Paul M. Barrett, Rodolfo A. Coria, Jean Le Loueff, Xu Xing, Zhao Xijin, Ashok Sahni, Elizabeth M. P. Gomani, and Christopher N. Noto. Dinosaur distribution // / Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). — 2nd ed. — Berkeley: University of California Press, 2004. — P. —606. — ISBN 0-520-24209-2 .
|
|
|
|---|


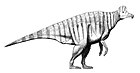
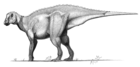
- 2021-09-06
- 1
