Interested Article - Синокаллиоптерикс

- 2021-06-21
- 1
Синокаллиопте́рикс ( лат. Sinocalliopteryx ) — род тероподовых динозавров из семейства компсогнатид (Compsognathidae), окаменелые остатки которого найдены в нижнемеловых отложениях формации (129,4—122,46 млн лет назад).
Известны два экземпляра. На обоих образцах сохранились отпечатки нитевидных «протоперьев», характерных для многих тероподов формации Исянь. Образец голотипа сохранил часть ноги дромеозаврида в области пищеварительной системы , а также возможные гастролиты . Второй известный образец показывает следы двух приёмов пищи, в том числе кости конфуциусорниса и лопатку птицетазового динозавра .
Описание
Синокаллиоптерикс был мелким бипедальным хищником . Сохранившаяся длина голотипа составляет 237 см . В 2016 году оценил массу тела этого динозавра в 20 кг . Синокаллиоптерикс отличается от родственного ему таксона Huaxiagnathus относительно длинными передними конечностями. Он является самым крупным известным компсогнатидом (которые сами по себе были довольно мелкими), и, в связи с размером тела, его передние и задние конечности были пропорционально длиннее, чем у прочих компсогнатид .
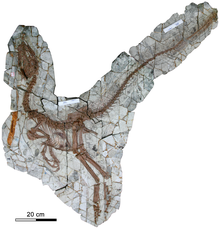
У синокаллиоптерикса была вытянутая голова с заострённой мордой и выпуклым верхним профилем. Предчелюстная кость несёт 4 мелких зуба с пилообразной насечкой на переднем крае. Верхнечелюстная кость голотипа содержит только 6 более крупных зубов; при жизни животного зубов было больше, но точное их количество определить не представляется возможным. Другой известный образец, под инвентарным номером CAGS-IG-T1, сохранил 10 верхнечелюстных зубных лунок с местом для 1 или 2 зубов в повреждённых местах. Скуловая кость необычайно крепкая, с высокой передней ветвью, которая являлась частью нижнего переднего края глазницы . Нижняя челюсть не имеет отверстия на внешней стороне .
Позвоночник состоит из 11 шейных , 12 грудных , 5 крестцовых и как минимум 49 хвостовых позвонков . Конец хвоста отсутствует. Невральные дуги и шевроны хвоста сильно наклонены к телам позвонков. Брюшные рёбра имеют очень короткие боковые сегменты .
Передняя конечность имеет короткую плечевую кость , с локтевой костью , на задней части которой имеется слабо развитый локтевой отросток. Кисть сильно удлинена — её длина равна длине плеча и предплечья вместе взятых. Вершина II метакарпалии расширена в сторону I метакарпалии, что делает пястные кости более компактными. Коготь II пальца вытянут, достигая длины I пальца с когтем. III метакарпалия довольно короткая и несёт тонкий III палец .
В тазу имеется небольшая выемка на переднем крае подвздошной кости . Относительно длинный вертел седалищной кости изогнут вниз. Задняя конечность удлинена, в основном из-за вытянутой голени , имеющей 90 % длины бедренной кости . Нога животного также была длинной, особенно плюсневые кости .
Как и многие другие тероподы формации Исянь, синокаллиоптерикс сохранил «протоперья» — простые нитевидные образования, покрывавшие кожу и имевшие сходство с перьями у синозавроптерикса . Длина покровов синокаллиоптерикса различна — самые длинные «протоперья» покрывали заднюю часть бёдер и основание хвоста. Их длина доходила до 10 см . Они были обнаружены и на верхней части стопы . Хотя эти образования не были такими же длинными и не выглядели настолько современными, как соответствующие перья «четырёхкрылых динозавров», таких как Microraptor и Pedopenna , они указывают на то, что перья на ногах или их аналоги возникли у гораздо более «примитивных» ( базальных ) динозавров, чем ранее известные .
Открытие и название

Типовой вид Sinocalliopteryx gigas назвала и описала в 2007 году группа китайских палеонтологов под руководством Цзи Шуаня и Цзи Цяна. Родовое название динозавра происходит от лат. Sinae — «китайский», греч. καλός/kalos — «красивый» — и греч. πτέρυξ/pteryx — «перо». Крупный (для компсогнатида) размер динозавра нашёл отражение в его видовом названии, которое переводится как «гигант» .
Голотип JMP-V-05-8-01 был найден в китайской провинции Ляонин , в стратиграфическом горизонте Цзяньшангоу формации Исянь, который датируют барремским — аптским ярусами нижнемеловой системы (129,4—122,46 млн лет назад). Он принадлежит взрослой особи и состоит из почти полного скелета с черепом, сжатых на одной плите. Образец сохранил чёткие отпечатки нитевидных «протоперьев» . В 2012 году был описан второй образец, получивший инвентарный номер CAGS-IG-T1 и принадлежавший животному более крупному, чем голотип. Его череп был приблизительно на 10 % длиннее, ноги — примерно на треть; разница объясняется положительной аллометрией . Образец состоит из частичного черепа, хвоста, конечностей и грудной клетки. Как и голотип, он сохранил отпечатки нитевидных образований .
Систематика
Авторы описания уверенно отнесли синокаллиоптерикса к семейству компсогнатид . Одним из диагностирующих признаков этого семейства является сходство морфологии метакарпалий (пястных костей). В работе 2007 года проводились исследования морфологии I метакарпалии нескольких видов компсогнатид. Авторы исследования пришли к выводу, что представители группы обладали явно выраженной морфологией кисти, где, как свойственно тероподам, первый палец больше двух других, но с отличающимся строением I метакарпалии, более короткой и толстой. Кисть компсогнатид также имеет выступ, который располагается на этой метакарпалии .
Приведённая ниже кладограмма показывает филогенетические связи синокаллиоптерикса внутри семейства компсогнатид согласно исследованию и Симоне Маганучо (2011) .
| Tyrannoraptora |
|
||||||||||||||||||||||||||||||||||||||||||||||||
Крупный размер синокаллиоптерикса по сравнению с другими представителями компсогнатид может указывать на тенденцию к увеличению тела внутри этого семейства — группы, известной своими скромными размерами по сравнению с гигантскими тероподами. Эта тенденция, очевидно, является общей для прочих линий динозавров .
Палеоэкология

В пищеварительном тракте хорошо сохранившегося экземпляра голотипа синокаллиоптерикса учёные обнаружили остатки ноги дромеозаврида , включая целую голень и стопу с пальцами и когтями в их естественном, сочленённом состоянии . Часть ноги довольно велика по сравнению с брюшной полостью — около 30 см , но она определённо располагается внутри неё, между рёбрами . Цзи и его коллеги в 2007 году пришли к выводу: находка может указывать на то, что синокаллиоптерикс охотился на более мелкого птицеподобного динозавра. Открытие показало, что он был активным и ловким хищником; такой вывод подтверждают находки других компсогнатид с остатками ящериц и мелких млекопитающих в брюшных полостях . В исследовании 2012 года нога дромеозаврида была предварительно идентифицирована как принадлежавшая особи синорнитозавра длиной 1,2 метра . В той же работе сообщалось о недавно обнаруженных остатках пищи в брюшных полостях и других компсогнатид (к примеру, окаменелые кости рыбы и лепидозавров были найдены в хорошо сохранившемся пищеварительном тракте сципионикса ). Чуть выше остатков ноги дромеозаврида видны отпечатки перьев. Ниже можно увидеть два кластера переваренной пищи. Было высказано предположение, что перья принадлежали птице и попали в кишечник отдельно от ноги, которая всё ещё оставалась там. Исследователи, сравнив находку с сохранившимся отпечатком пищеварительного тракта сципионикса, определили, что на момент смерти синокаллиоптерикса переваренная пища размещалась в его двенадцатиперстной кишке . С-образное содержимое брюшной полости в этом образце отражает первоначальный контур пищеварительного тракта .

В дополнение к дромеозавридовой ноге в брюшной полости были обнаружены 4 камня неправильной формы диаметром от 15 до 20 мм , в других частях скелета или в окружающих отложениях подобные камни отсутствовали. Авторы описания интерпретировали их как гастролиты , сходные с найденными у Nqwebasaurus и барионикса . Другие тероподы, такие как каудиптерикс и неназванный пока орнитомимид , также были обнаружены с гастролитами, хотя в их случае камни были более многочисленными и меньшими по размеру. Цзи и его коллеги предположили, что, поскольку два последних динозавра были, вероятно, в основном растительноядными, количество и размер гастролитов могли соответствовать рациону: растительноядные животные глотали много мелких камней, а хищники — меньшее количество более крупных, для улучшения пищеварения . Тем не менее исследование 2012 года не нашло гастролитов во втором образце, и потому палеонтологи пришли к выводу, что камни были проглочены животным случайно. В любом случае голотип не сохранил остатки мускульного желудка .
Второй образец синокаллиоптерикса, CAGS-IG-T1, также содержит следы двух приёмов пищи. Перед нижней седалищной костью были обнаружены раздробленные кости, принадлежавшие по крайней мере двум особям Confuciusornis sanctus — базальной птицы, которая была широко распространена в формации Исянь. Помимо этого, была обнаружена лопатка длиной 135 мм , принадлежавшая 1,5-метровому растительноядному птицетазовому динозавру, вероятно, Yueosaurus или одному из видов пситтакозавра . Поверхность лопатки выглядит так, как будто она была протравлена кислотой желудка в течение как минимум 13 дней ; это навело исследователей на мысль, что синокаллиоптерикс проглотил птиц позднее и сразу несколько. Это указывает на высокий метаболизм динозавра, который требовал регулярных приёмов пищи .
Тот факт, что второй найденный синокаллиоптерикс поймал двух птиц за короткий промежуток времени, а также присутствие перьев в его пищеварительной системе может указывать на то, что данный род динозавров специализировался на подобной добыче. Даже синорнитозавр мог оказаться его жертвой потому, что, как предполагают учёные, был летающим животным. В 2011 году был обнаружен образец микрораптора с остатками птицы в брюшной полости, что рассматривалось как доказательство древесного образа жизни микрораптора. И хотя синокаллиоптерикс был, по всей видимости, наземным динозавром, он вполне мог внезапно нападать на птиц — этот способ охоты используют многие современные животные .
Примечания
- Хоун Д. Хроники тираннозавра: Биология и эволюция самого известного хищника в мире / науч. ред. А. Аверьянов . — М. : Альпина нон-фикшн , 2017. — С. 180. — 358 с. — ISBN 978-5-91671-744-0 .
- . The Princeton Field Guide to Dinosaurs. — 2nd ed. — Princeton: Princeton University Press , 2016. — P. 124. — 416 p. — ISBN 978-0-691-13720-9 .
- ↑ Ji S., Ji Q., Lu J., Yuan C. (англ.) // Acta Geologica Sinica. — 2007. — Vol. 81 , no. 1 . — P. 8—15 .
- ↑ Lida Xing, Phil R. Bell, W. Scott Persons IV, Shuan Ji, Tetsuto Miyashita, Michael E. Burns, Qiang Ji, Philip J. Currie. (англ.) // PLoS ONE . — 2012. — August ( vol. 7 , no. 8 ). — P. e44012 . — doi : . 13 октября 2018 года.
- Gishlick A. D., . On the manual morphology of Compsognathus longipes and its bearing on the diagnosis of Compsognathidae (англ.) // . — 2007. — Vol. 149 , no. 4 . — P. 569—581 . — doi : .
- ↑ ., Maganuco S. (англ.) // Memorie della Società Italiana de Scienze Naturali e del Museo Civico di Storia Naturale di Milano. — 2011. — Vol. XXXVII , no. I . — P. 123 . — doi : .
- Smithwick F. M., Nicholls R., Cuthill I. C., Vinther J. (англ.) // Current Biology . — 2017. — Vol. 27 , no. 21 . — P. 3337—3343.e2 . — doi : . 31 мая 2019 года.
- 2021-06-21
- 1
