




Млекопитающие неогена
- 1 year ago
- 0
- 0

?????????????? ( ???. Mammalia ) — ????? ??????????? ???????? , ???????? ????????????? ???????????? ??????? ???????? ????????????? ????????? ??????? . ????? ?????? ? ????? ???????? ????????? ???????????? ? ???????????? ????? ????? ????????????? ?????? ??????????? . ?? ????????? ????????, ???????? ????? 6500 ??????????? ????? ????????????? , ????? ??????? ? ??? ??????? ???????? ( ???. Homo sapiens ); ????? ?? ????? ????? ?????????????, ??????? ????????, ????????? 20 000 .
????????????? ?????????????? ????? ??????????? . ????????? ?? ?????????????? ???????? ? ????? ?????? . ? ????????????? ??????? ???? ??????? ????????????? , ???????? ? ???????????? , ? ????? ??????? ????? ???????? ?????. ??????????? ????, ???????? ? ????? ?????????????, ?????????? ? 26—29 ??????? .
??????? ??? ????????????? ?????? ????????????, ?? ? ????? ????????????? ???????????? ? ??? ?????? ???????????? ????? ???????? . ???????? ? ?????????? ????????????? ??????????????? ???????? ??? ?? ?????????????? ??????, ??? ? ? ?????? ????????????. ?????? ?????? ?? ???? ?????? ???????? ???????? ?????? ????????, ??? ??? ?????? ????? ????? ??????????? ??????? ???????? ???????????????????? . ?????????????? ????????????? ? ????? ? ????????? ?????? ???????? ?????? ????????????, ?? ????????? ?????????? ?????????? ? ????????????? . ??? ???????? ???????????? ??????? ????? ???????? ????? (? ?????? ????? ??? ???????? ???? ????????? ????? ). ????????????? ?????? ??????? ???? ? ????? ? ????????????? ???????????? ????????: ??? ????????? ??? ?????? ????????? ????????? ??????? ? ????????????????? ????? , ????????? ???????????? ???????, ?????? ???????? ?????, ???????????? ??? ???????????? ???????? ? ???????? ??????? .
????????? ????????????? ?????????? ????? ?????????? ( ????????????? ) .
????????? ???????????? Mammalia ???????????? ????? ??????????? ?? ???. mamma — « ????? , ???? » (???? ? ????? ? ???????? ???????? ????? ? ???? ?????????????) . ????? ?? ????? ????? ??. mammifères , ???. mamíferos , ????. mammiferi , ????. mamíferos , ?????. mamuloj ( ????. mammals — ????????????? ?? ??????). ???????? «?????????????? ???????» ????? ???????? ???. Säugetiere , ??????. zoogdieren , ???. ????????????? ; ???????? «??????? ??????» — ???????? ??????. ssaki , ???. .
????????????? ?????????????? ?? ????? ???? ? ??????????? ?? ???? ??????????? , ?? ???? ??????? ? ?? ??????????? ???????????? ???????? . ??????????? ????? ????????????? ?????????????? ?????????? , ????????? ? ????? ??????? , ???????? — ??????????? ?????????, ??????? ? ????? ???????????? ??????????? . ????? ??????? ??????????????? — ? ???????????? , ??????? ??? ? ????? ??????? ????? ????????????? ????????? ???????????? ?? ???? ???????????, ????? ????????? ? ?????????? , ?????????? ?? ??????????? ??????, ???????? ? ????????? ? ??????? ????? . ? ????????? ?? ?? ?????? ??????????? ??????????? ???????, ?? ?????? ???????? , ???? ???????? ???? ????????????, ? ?????? ??????? ???? ? ??????? . ?? ?????????? ?? ?????? ????????? ???????? ?? ???????? ???????? ???????????? ???? ?????? ?????? ????? ?????????????: ?? ?????? ?? ???, ? ??? ????? ? ? ????? ???????? , ?? ???????????? ?????? ????????? ????? ??????? ????? .
????????????? ???????? ????? ??? ??????? ????? ? ??????????? ??? ? ???????? ? ??????????? ????? , ??? ? ? ??????????? ? ? ???????? ????????. ? ???????? ???????? ? ????????, ? ???????, ?? ??????????? ??????? ?????????? ?????, ??????????? ?????????????, ????????? ????????????? ????? ? ?????????????? ????????? , ???? ? ??? ????????? ??????????? ?????? ? ???? . ?????? ? ???????, ????? ?????????? ??? ?? ???? ?????? ??????? ??????? ????? , ????????????? ???????? ? ???? ???????: ?? ???? ??????????????? ??????????? ? ?????? ?????????? ??????? ?????????????? , ???????? ( , ????????????? ) ? ???????????? ( ???????????? , ?????????? , ???????????? ) . ????????? ??????????? ???????? , ? XX ???? ??????? ?????? ? ?????, ??????????? ??? ?????? ?????????????: ?????? ?????? ?????? , ???????? ?? ??????? ??????????? ? ?????? ?????????? ??? , ????????? ?? ??? ?????????? ??????? , ????????? ? ???????? ???????????? ???????????? . ???????, ? ????????? ?? ???? ??????????? ???????? ????????? ??????? ?????? : ?. ???????? ???????? ?? ?????? ?????? ?? ???????? ????????, ? ????? ????????? ???? ?? 3,5 ???? ?? ?. ?. ???????? .
? ?????? ????????????? ????? ???????, ????????? ???????? ?? ?????????? ???????????? ????????????? ? ???????? ????? (????????????? ?????, ??????? ???????, ????????-????????? ? ?????????????? ???????) ? ??????? ???????????? ??????????? — ????????, ??????? ??????????? . ???, ??????????? ????????? , ????????? ? ????????? ? ????????? ????????? , ????? ?????? ? ??????? ??????, ???????? ??????? ; ????? ?????????????? ????????? ???????????? ?????? ??????????? ? ???????? ???????? . ???? ????????? ???????? ??????? ????????????? ?????? ?????? ???? ??? ????????????????? ????????????? ????? . ?????????? ?????????? ?????????? ????? ???????? ? ????????? ???????: ????????? (201 ??????? ?? 436 ????? ??????? ??????????), ????????? (198 ?? 252), ??????? (140 ?? 450), ??? (101 ?? 428), ????????? (97 ?? 153), ???????? (96 ?? 394) .
????? ????????????? ????????? ??????? , ??? ??????????? ????????????? ?????????? ????? ??? ?????????, ?????????? ??, ??? ???????, ?? ?????? ??????????? :

???????? ??????????? ??????????????, ????????????? ????????? ?? ????? ??????????? ???????? ?? ?????? ?????????? , ??????????? ? ????? ?????????? ??????? . ???????? ??????????? ???????????? ????????? ??? ?????? ?????????? ????????????? — ???, ????????, ???????????? ?? ??????? Oligokyphus ?? ????????? Tritylodontidae ? ??? ???????? ???????? ????????, ?????? ? ??????? ?????? ? ?????? ??? . ????? ?? ????? ????? ? ????????? ??????????? ?????????????: ? ?????????? ???????? ?????? ??????? ?????????? ??????? ?????????????? , ??????????????? , ? ???????? . ????????? ?????? ????????????? ??? ?????? ?????????????? ????????? (??? ???????????) ?????????? , ???? ???????? ????? ??????????????? — ???????? ????????????? ? ?????????????? ?? ??????????? ??????? ?????????????, ????????????????? ????? 100 ??? ??? ; ??? ???????? ???????????????, ?? ??? ?? ?????? ?????? ? ???????? ??????????? ?????? ? ??????????????? ?????? ???? ????? ??????? ????????????? .
? ??????? ?????? ?????????? ? ?????? ???????? ????? ?????????????, ????????? ??????? ??????? ????????? ? ????? ???????? ???????: ?????, ?????????? ????????????? ; ????? ????????????? (??? — ??? ) ; ???????, ?? ?????, ? ??????? ??????????? ???????? ? ???????????? , ???????????? ???? ?? ????? ? ?????? ??????? .

??????????? ???? ????????????? ???????? ?????? ?????????? ??????. ??? ?????? ? ???? ??????? ? 1758 ???? ??? ????? Mammalia ?. ??????? ? 10-? ??????? ??? « ??????? ??????? » . ? ?????? ??????? ?????? ???????? ????? 184 ???? ????????????? ; ? ????????? ?? ????? ??????????? ???? ????????????? ?????????? ? 26—29 ??????? , 153 ????????? ? 1229 ????? .
??????? ? ?????? ?????? ????????????? ???? ???????? ?????????????? ???????? ??????; ????????????? ????????? ????????? ?? ??? (????????, , ????????????? , ???????????? ? ????????? ?????? ???????? ? XIX—XX ????? ??????????? ????????) ? ?????? ????? ???????? ???????? ? ???? ????????????? .
???????????? ????????????? ?? ?????? ???????????? ? ????, ???????????? ? ????? XIX ????, ???????? ????????? ??????? :
????? Mammalia Linnaeus, 1758

????????, ??????????? ? ??????, ??????????? ???? ??? ?????????????? ???????????? ??????????????? ???????????? , ???? ??? ???????? ?? ????????? ????? .
??????????? ??????? ?????? ????????????? ?? ?????- , ????- ? ???????? ???? ?????????? ??? ??????????? ???????? ?. ????????? (1839), ??????? ??????????? ??????? ?????????????? (??????????????: «?????????????», «???????????», «????????????») .
???????????? ???????????? ????? ??????????? ????????????? ?????????????, ???????????, ? ?????????, ????????? ???????? ??????????????? ????? ??????????????? ????????? ????????, ?? ????????????? ?? ???? ?? ??? ?? ????? .

?? ??????? ???? ???????? ??????? Theria , ??????????? ????????????? ?????????????, ?????? ??????????? ??? « ????? » (? ?????? ???????? ??? ????????????? ??????????? ?? ?????? «?????»; ??. ????? ). ?????? ?? ????? «?????» ?????????? ??? ??????? ????? «?????????????» . ???? ??????? ??????? ? ????????? ?????????????? ????????: ???? ??? ???????? ?????????????, ??????????????????? ????????? Prototheria ( ?????????? ) , ???? — ????? ?????????? ?? ???? ???? ???????? ????? — ?????? ??????????? ??????? ????? (? ??????? ???????? — ???? ?? ????? ???? «??????????» ).
????????? ????????????? ?? ????????????? ???????:
|
??????????? ??????
(Mammalia)
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
|
|||||||||
|
Prototheria
(??????????) |
|||||||||
|
Metatheria
(?????????) |
|
||||||||
|
Eutheria
(???????) Placentalia (????????????) |
|
||||||||
?? ???????? ? ???????? ?????? ????????????? ????? ????????????. ???? ? ?? ???????????? ??????????? ????, ??? ? ? ?????? ?????? , ????? ?????????? ?? ?????? , ??? , ???????? , ??? ???? ??????????? ? ????? , ????? ? ??????????? ???? ?????? ???? ? ?????? ????? ?????????, ??????? ?????????????? ? ????? ???????? ? ?????????????? ????????? ???????? . ?????? ???????? ??? ????????????????? ??????? ??? ?????? , ??????????? ??? ??????? ?????? ? ??????? ?????? ? ???? ???????? , ??? ? ?????? ?????????? ??????? ????????? ? ???????????? . ?????? ????????? ? ????????????? ????? ??????: ???? ? ?????????????? ????? ( Manis tetradactyla ) ? ??????????? ?????????? ( Euchoreutes naso ) ?? ? ??? ???? ??????? ????????, ?? ? ??????? , ??????? ???? ? ???? ?????????? ????? ????????? ??????????? . ????? ??????, ? ????????? ??????????? ??? ??????? ? ???????? ??????, ??????? ?? ??????????? ????. ???, ????? ??????? ??????? ????? ? ?????? ? ???????? , ??????? ????????? ??????? ???? ?????? ???????? ?, ??????????????, ??????? ????? ?????? ?????????????. ????????, ??????? ???????????? ?????????? ? ?????? ( ?????? ), ???????????????? ( ??????? ), ????????? ?????????? ??????????? ??????????? ????, ? ????????????? ??????? ?????? ?????? ( ?????? ???? ) .
???????? ?????? ?? ???????? ?????? ?? ?????? ????????????? ?????????? ?????????? , ? ??????? ???????? ?????????? ?????????????? ? , ? ????? ?????? ? ???????????? , ???????? ?????????? ??????? ????? ???????? ? ??????? ?????? ????? ???????????? ? ???????? , ? ?????? ??????? . ??????? ? ???????? ??????????? ? ?????? ??????? ????????????? ?????????????? ?????????? ? ???? ?? ?????????? ? ?????? ?????? «?????» . ?????? ? 1693 ?. ??. ??? ?? ???? ??????????????? ??????? ?????????? ? ????? ? ?????? ??? ???????????, ??????? ???? ???????? «??????????????» .
?????? ??????????? ????????????? ????? ?? ????? ? ???????? ????. ?????????? ???????????:

|

|

|

|

|

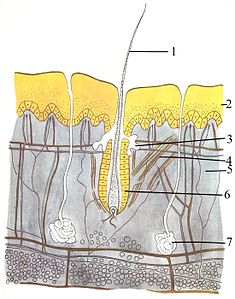
???? ????????????? ?????????? ???????????? ???????? ? ??????? ????????? ?????? ????? ? ??????? ???????????. ??? ? ? ?????? ??????????? , ??? ??????? ?? ???? ?????: ???????? ??????????????? ( ????????? ) ? ??????? ??????????????? ( ????? ) .
??? ????????????? ?????? ?????????? ??????? ??????????? ?????????????? ?????????????: ??????, ????? , ????? , ?????? , ???? ? ??????? . ??? ???? ?????? — ???????????, ???????? ????????????? ????????????? ?, ??-????????, ???????? ??????????? ?????????? ; ??? ?? ??????????? ?????-???? ??????????? ???? ?????? ??????. ??????? ??????? ????? — ?????????????, ???? ??????? ??? ????????? ?????????? ??????? ?????????, ?????? ?????, ???????????? ??????? . ?????? ???????????????? ? ???????????? ???????, ?????? ???????? ???? ?? ??????????? ? ????????? , ???????? ????- ? ????????????????? ???????? ????, ???????????? ????????????????? ???????. ???????????? ???????? ??? ?????? ?????????? ????? ? ???? ????????????? ( ????? , ?????? , ????????? ???????? , ??????????? , ???????????? , ???? ) ????? ????????? ???????? .
? ??????????? ????????????? ???????? ??????? ??????? ???????? ??????? , ??????????????? ??????? ???????? ??????? ????? , ????????? ? ?????? ???????? , ? ?????? , ??????????? ??? ???????? (????? ??? ?????????? ???????????? ). ?????? ????????????? ????? ?? ?????? ??? ??????????? ??????? ??????? , ? ? ?????????? ? ??????????? ??????? ??????? ????? , ?????? ???????????? ????????? ? ?????????????? , ????????? ??? ???? . ??? ???? ????????????? ( ???????????? , ???????? , ??????????? , ????????? ) ?????????? ??????? ?????? «????????????», ??? ???? ; ?????? ??? — ????????? ??????????? ???????????? ???????? , ???????????? ??? ?????? ? ????????? . ???? ?????????????? ????????? ?????? ?????????? ???? ?????????? ?? ????????? , ? ??????? ??????

??????????? ??????? ???????? ?????? ??????? ???????? , ?????????? ?????????????? ??? ?????? ????? ???????? ??????? ?? ??????????? ???? ??? ????????? ???????? ?????????? ??????? (????????, ??????? ????? ????? ????????? ??????? ?????? , ?????? ? ????????????? ?????? , ??????? ???????, ??????? ? ??????????????? ?????, ?????? ?????? ???? ? ???? ??????????? ) .
??????????? ???????????? ????????????? ???????? ??????? ???????? ?????? ????? , ??????? ??? ???????????? ????? ????? ??????????? ??????? ?? ??????????? . ? ?? ????? ????????? ??????? ? ??????? ?????? , ???????? ??????? ???? ??????? ? ?????????? ?????????? ??????? . ??????????????? ??????? ????? ?? ?????? ????????????? ???????? ??????? ?????? , ??????? ? ???????? ??????? ???????? ?????? — ???????? ???? ????????? ?????????????. ? ?????????????? ???????? ? ???????????? ??????? ??????? ????? ??????????? ?? ?????? (??????? ?????? ????????????? ?????? ?? ?????). ????? ?????? ????? ??????????? ? ?????? ????? ?? 2 ?? 24, ?????? ? ?????? ???????? ????? ????????? ? ?????? ???? .
??????? ?????? ??????????? , ????????????? ????? ?????????? ?????? (? ??????????? ???? ? ??????? ?????? ? ???? ??????? — ??? ?????????? ? ???????????). ?????? ???????? ????? , ??????????? , ??????? ?????? , ????? ??????? ? ?????? ??????????? ? ???? ?????????? .
????? ????????????? ????? ???????????? ??????? ???????? ???????, ??? ? ???????????????? ????????????? ??? ? 3—4 ???? ???????, ??? ? ???????? ???? ?? ??????? . ??-?? ????????? ????????? ?????? ????? ????? ????? ????????? ?????? ? ?????? ??????, ??? ? ?????? ??????????? . ? ????????????? ??????????? , ? ???????? ????????? . ?? ??????????? ?????? ? ???????? ?????? ?? ?????? ???????????? ????????? ??????? ??? ???????? ?????? , ??????? ?????? ???????? ? ?????? ???????????? ???????, ???????? ???????? ? ?????? . ??? ???????? ??????? ????????????? ?????????? ??????? ???????? ???? , ???????????? ???????? ?????? ? ???????? ????????? ?????????? ????? (??? ?????? ???????? ?????????? ???????? ? ?????????? ????? ???????? ????? : ????????? ??????????? ??? ?????????? ?????? ????? ????????? ??????????, ?????????? ? ?????? ), ? ??????????? ?????? ? ??????? ??????? ???????? ( ??????? ) ??? ?????? ???? ???????? , ???????????? , ???? ????????????? . ??????? ??????? ?????????? ???????????????? ? ????????????? ???????; ?????? ?? ??? ??????????? ??????? ????. ?????? ???????? ??????????????? ?????? ?????? ? ??????? ??????? ???????? ????? ??????????? ??? ????????????? ????????? ??????? ???? , ? ?????? ??????? ? ??? ???????????? ???? ??????? , ??????????? ?????? ???????? ? ?????????? ?????? ???????? ????. ???????? ??????? ????????? ???????? . ?????? ????????????? ??????????? ????????????? ? ??? ?????? ?????? ?????????? ?????; ????????? ? ?????? ???????? ? ???????????? ???????? ??????????? ? ?????????? ????? ?????????? ? ????????????? ?????????? ???????? ??? — ?????????? ? ??????????? .
??????????? ????? ? ???????? ???????????? ?????? ?????????????: ??????? ??????????? ????????? ( ???????????? ???????? ), ????? ???????? ????????????? ???????? ?????; ???????? ??????????? ???????????? ?? ???? ??????? — ??????, ???????, ??????????, ?????????? ? ????????? ( ??????? ??? ???? ? ??????????? ???????????? ); ?????????? ????? ?????? ????????? — 7 (??? ??????????, ??? ????? 6 ? ????????? , ? ? ????????? ???????? ????? ?????? ????????? ?????????: ? ??????? ???????? — ?? 5 ?? 6, ? ????????? ???????? — ?? 8 ?? 10 ) . ??????? ????? ???????????? ? ????????????? ???? ????? ??????? ?? 12—15, ?????????? — ?? 2—9, ?????????? — ?? 1—9 ?????????; ??????? ????? ????????? ????? ????????? ?????????: ?? 3 ? ?????????? ?? 49 ? ?????????????? ????? . ? ???????? ??????? ????????? ???????????? ????? , ??????????? ? ???????? ? ?????????? ?????? ? ??? ??????? ?????? .

???? ???????? ??????????? ????????????? ?????? ??????? ?? ???? ??????? ? ???? ?????? ; ?????????, ???????, ??????? ???????????? ??? ????? ???????????. ? ????????????? ??????? ??? ???????? , ??????? ? ????????? ????????????? ???????????? ? ?????????? ? ??????? . ? ???????? ?????? ?????? ?? ???????????? ? ????? ????? ???????? ??????????? ??????? ?? ????? , ?????????? ? ????? ; ????????? ???????? ????????, ????? ? ??????, ??????? ?????? ??????? ?? ????-???? ?????? (?????? ? ???????????? ????? ?????? ????????? ? ????????? 13—14). ???? ?????? ??????????? ? ?????????????, ??? ???????, ??????? ?? ?????? ???????? , ?????????? ? ??????????? ??????; ? ?????? ?????? ?? ?????? ??????????? ?????? ????? , ?????? ? ????? (?????????? ?????????? , ?????? ? ????? ??????? ). ? ??????????? ?? ?????? ????? ?????????? ????????????? ???????????? ????????? ??????????????: ? ???????? ???? ??????????? ?????????? ????? ? ??????, ? ? ?????? ???? ??? ?????????, ?? ???????? ??????? ??????? .
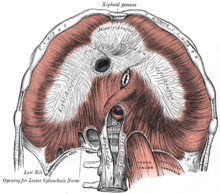
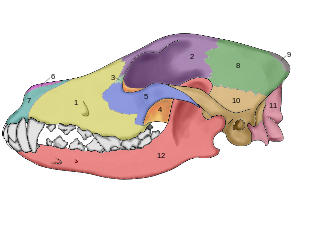
???????????????????????? ???????? ??????? ????????????? ???????????? ?????????? ????????????? ???????? ? ????? ? ?????? ??????? ? ?????????? ???????????? ??????????? ? ??????????? ?? ??????????? ???????? ????????????. ????? ????????? ???? , ?????????? ???????? ???????? ????? ??????????? ; ??????????? ??????????? ??? ???? (? ???????? ? ?????? ???????? ??? ????????????? ?? ????? ????) ????????? ???????? ???? ? ????????????? ?????? ????. ??????????? ????????????? ????????? ????????????? ???????? ????? ??????? ?????????????? ????? — ????????? , ??????? ????????? ??????? ? ??????? ???????; ?? ????????????? ????????? ????? ????????????????? ?????????? ?????? .
? ????????????? ?????? ??????? ??????????? ??????????? , ?????????????? ?????? ? ???????????? ????????? ????, ? ????????? ??????????? , ??????????? ? ?????????????? (????????? ????????? ?????????? ???????) ? ??????? ???????? (???????? ?? ?????? , ???????? ???????? ? ?????? ? ???????? ). ??? ????????????? ???? ???? ????????? ? ?????? ?????????-????????? ; ????? ?? ?????????? ??????? ????????, ??? ???????, ???????? . ?????? ????????? ???????? ????? ??????? ? ?????? (????????? ??. ?????? « »).
??????? ????? ?????????????, ??? ? ? ?????? ??????????? , ??????????????? ????????????? ? ??????? ?? ??????? ?????? ( ???????? ) ? ??????????????? ?????? — ????????? . ??????? ???????? ?????????????? ???????? ( ??????? ) ???? ? ?????? ? ???????????? ????????????? ? ??????????????? ??????? ?????????; ????? ???????, ?????? — ??????????? ? ?????????????? ??????? ??????? ??????? . ???? ?? ??????? ????? ???????????? ?? ????? ???????? , ???????????? ?????? ???????? ? ???????????????????? ???????? ????????? , ? ????? ???????? , ???????????? ?????????????????? ???????? ????????? . ??????? ??????? ???????????????? ?? ??????? ???????? ? ?????????? ?? ??????? ? ??????? ??????????? ??????? ???? ???????????? ????????? — ???????? . ????? ? ???? ????????????? ????????????? ???????? ?????????????? ? ??????? ?????????? ?????????? ????? ?????? ??????? — ??????????????? , ? ?? ????????????? ???????? .
??? ? ? ?????? ???????????, ??????? ??????? ????????????? ?? ??????????????? ????????? ????? ?? ??????????? , ?????????? ???????? ? ??????? ???? , ? ?????????????? , ??????? ????????? ?????????? ?? ????????? ? ???????? ????? ??????? .
? ??????????-?????????????? ????? ??????? ??????? ???????????? ?? ???????????? , ????????????? ????????? ??????????? ? ?????? ?????? , ? ???????????? , ????????????? ?????????? ?????? , ??????????? ? ????????????? ?????? ; ??????? ????????????? ????? ??????? ????????? ?? ??????????? .
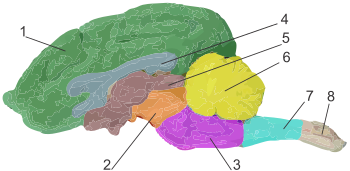
????????????? ??????? ????????? ????? ? ????????????? ????? ??????: ?? ????? ?? ? 3—15 ??? ??????????? ??????? ???? (? ???????? ?? ????? ???????? ?????????) . ???????? ???? ????????????? ???????? ???? ???????: ???????? (??? ???????? ), ????????????? , ??????? , ????????????? ???? ? ???????? , ?? ??? ???????? ??????? ?????? ? ????????? . ??????????? ???? ????????? ?????????? ????????? ???????? ???? ( ?????????? ), ????????? ????? ???? ???? .
??? ???????? ????? ????????????? ?????????? ???????????? ???????? ????????? ?????? ???????? , ??? ??????? ??? ???????? ? ?????????? ??????? ?????? «???????» ? ???????? ????????? — «??????». ? ????????????? ??????? ?????? ?????????? ???? , ?????? ?? ???????????? ???? ???? ??????? ????????? ????? ? ??????????? ???????? ?????????????? ? ???????? ????? .
?????????????? ??????? ??????? ????????????? ???????? ???????? ? ?????, ????????? ?????????????? ?? ????????? ? ???????? ?????, ? ????? ?? ??????????? (????? ? ????????? ?????????????? ??????????? ????? 200 ?????? ?????? ? ?? ?????????????). ? ?????????????, ??? ? ? ?????? ?????? , ??????? 12 ??? ???????? ??????; ????? ?????????????? ?????? ??????????? ?????? (? ???????? — 31 ????) .
??????????? (11 ??? ?? 12) ???????? ? ????? ?????????????? ?????? ????????????? ??????? ? ???????????? ??????? ??????? , ??? ????? ??????????? ???? , ??????? , ????????? ????? ? ?????? ?????? . ?????? ????? ?????????????? ??????, ? ????? ? ???????????? ??????? ?????????? ???? ????????????? ??????? ? ???????????? ??????? ??????? . ?????????????? ? ????????????? ????????? ????????????? ? ????????????????? ???????????? ??????? ???????. ? ?????? ?????? ?? ??? ?????? ???????????? ?????? ? ??????? ????? . ??? ????????????????? ??????? ???????? ????????: ??????? ????? ???????? ?? ????????????? ??????? ??? ???? ? ?? ???????; ?????? ??????????? ? ?????????? ?????? ???????? ????? ? ? ???????? ????? . ??? ???? ???? ? ??? ????????????? ? ????????????????? ??????? ????? ?????? ????? ?????, ?? ? ???????????? (?, ? ?????????, ? ?????????????) ?? ??????? ?????????? ????????????? ???? ?? ????? ?????? ?? ??????? ????????? ???????????? .
?????? ?????? ? ????????????? ???????, ??? ???????, ?????????? ??????, ???? ? ????? ? ?? ????? ??????? ????????, ??? ? ???? . ?????? ????? ?????????? ?? ???????? ???? ????????? ????? ?????? . ????????? ?????????? ???? ????? ?? ???????? ?????, ? ?????????? ????????? ????? ?????????? ??? ???????? ?????, ??????????? ??????????? ????? . ? ??????? ?? ?????????????? ? ????, ? ????????????? ??? ??????????? ?????????? ????? ??????????? ? ????????? ?? ??????????, ? ? ?????? ???????? ( ??????? , ???? ) ??-?? ???????????????? ?????? ??????????? ? ??????????? ??????????? ???????? . ?????????? ????? ???????????? ??????????? ?? ???????? ????????????? : ??????? ???????? ?????? ?? ?????????? ?????, ? ???????? ???????????? ??????????????? . ???????, ???????? ?????? ? ????????????? ??????? ??????, ??? ? ????, ?????? ???????????????? (????????????????) ???????? ??? ????? ?????? ? ?????? ???????? , ? ? ???????????? ??????????? ????????????? ?????? — ??????????????? .

?????? ????? ? ???????????? ??????????? ????? ????????????? ?????? ??????? ? ??? ???? ???????? ????????????? ??????? . ? ?????? ??????? ?????? ????? ?????? ?????????? , ??????? ? ???????? ??? (?????????? ???????? ???????? ?????? ? ????? ???????? ). ???? ?????????? ? ??????? ??? ??????? ? ? ?????? ???????????? , ?? ???????? ??? — ???????????????? ?????????????, ??????????? ??????????? ???????? ???? . ????? ???????? ?????? ???????? ? ???????????? ??????????? ?????? (?? 6—8°) ??????????? ?????????? ????? (???????, ? ?????? ?????? ? ?????? ????? ????? ???????? ???????? ????????), ? ???????? ???????? ?????? ???????? ?? ??????? ??????????? ?????????? ????????? , ?????????? ??????? ???????? ??? ?? ????????? ??????? ? ???????????? ??? ???????????? ????????? ? ??? ?? ????????? ??????? ???????? ?????. ?????????? ??? ????????????? ? ??????? ? ??????? ???????? ??? ???? ???????? ???????? ( ????????? , ?????????? ? ????????? ) . ?????????? ???? ???????? — ??????? ????????? ????????? ? ???????? ?? ???????? ????????? ???? ??????????? ???. ????????? ?????????? ????? ???????????? ???????? , ??????? ? ????? ??????????? ???????? — ? ; ??? ???? ? ?????? ????????????? ????????? , ?????????????? ? ?????, ? ??????????? ??????, ??????? ? ??????? ?????? ? ?????? ?????????????? ???????? . ?????? ?????? ? ????????, ? ????????????? ????????? ?????? ?????????? ??????????????? , ??????? ????????? ?? ??????????? ? ????????? ??????????? ?????????? ??? ????????, ????????? ????????? ??????? ??? ?????????? ????????? ???? .
?????? ???????? ? ????????????? ??????? ???????, ??? ? ?????? ???????? ???????????. ??????????? ? ?????????? ??????? ????????????? ?????????? ??? ?????????????? ? ????????????, ??? ??????? ????, ? ?????? ?????????? ? ????????????? ?????????. ?? ??????? ???????? ???????????? ??????? ????????????? ???????????? ?? ??? ??????: ??????????? ? ????????????? ?????? ????????? (??????????? ?????????????) ? ??????????? ? ???????? ???????? ????????? ( ??????? , ???????????? , ?????????? ) . ?? ?????? ?????? ????????? ??????? , ? ???? ???????? ??????? ????? 5 ??? ???????????? ??????, ? ??????— ?????? , ? ??????? ?? ???????? ? 50 ??? ?????? . ? ????? ??????? ?????????? ???????? ??????????? ? ?????? ????? , ?????????? ????????? ???? ???????????? ??????-??????????: ? ??????????? ?? ?????? ????? 1000, ? ?????? ???????? — ????? 500, ? ???????? — ????? 387, ? ???????? — 262 . ??-????????, ???? ????? ??????? ???????? ? ????????????, ??????? ???????? ??????? ??????? ??????????? ???????????? ?????????? . ?????? ???????????????? ???????????? ??????? ?????????? ???????????? ?????? — ?????????? ??????? , ???????????? ????????? ???????? ? ????? ?? ?????????? ? ??????? ??????????? ?? ???????? ??????? ???????? ???????? .
?????? ????? ????????????? ???????????? ????????? ??????? , ??????? ??????????? ?? ????????? ????????? ????? ? ???????? ???? ? ???????? ????????? ????? (?????????, ??? ? ????????? ???????, ????????? ? ?????? ?????????????? ). ??????????? ?????????, ??? ??????? ?????????? ???????? ???????? ? ????????????? — ???????????????????, ?????? ?????????? ??????? ???????? ??????? , ??????? , ?????? ? ??????? . ???????????? ??????? ??????? ????? ??? ?????????? ?????, ?????????? ?? ?????????? « ??????? » ????? .
? ????????????? ?????? ??????? ?????????????????? ?????? ???????????????? , ? ??????? ??????? ??? ???? ????????? ??????????, ?????????? ?? ?????????? , ?????????????? ? ???? (? ?? ??????) ?????????. ????? ????? ?????????? ???????? ???????????????? (????????? ?????????? ???? ? ?????????), ??????????? (??????? ?????????) ? ????????????? ?????? ????????? ( ??????????????? , ?????????????? ) . ???????????? ???????????? ??????? ????????? ????? ??????? , ? ????? ???????? .
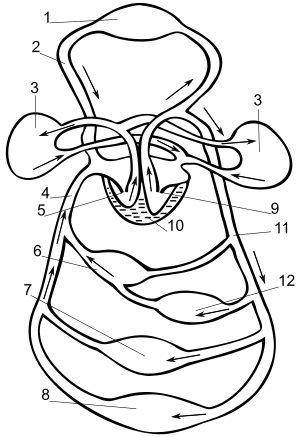
??????????? ??????? ?????? ??? ?????????? ????????? ? ?????????? ??????, ??????????? ??????????? ????????? ? ????????? ??????????? ???? . ? ????????????? ??? ???????????? ???????? ??????? ( ??????? ) ? ???????????? ?????? : ???????? ? ???????. ? ??????? ??????? ??????????? ????? ?????? ??????? ???? , ?????????? ? ?????????? , ? ???????? — ? ??????? ??????? , ????????? ??? ???? ????? ??????????? ? ???????? ???????. ??????? ?????? ??????????? ????? ??????? ?? ??????? (?????? ?? ??????? ? ??????? ??????????? ?????), ?????? ? ??????? .
???????? ??????? ? ????????????? ?????????: ??? ??? ?????????? ??????? ??????????? ????? ? ????????????? , ?? ???????????? ?????? ???????????, ? ????? ????????? ?????? — ?????? ??????? ????????? ????????, ???????? ??????? ????????? ???????? ????????? ????? .
? ????????????? ?????? ??????? ?????? ? ?????? (??????????? ? ??????????????). ? ???? ?????????????? ???????? ?????? ????????????? ????????, ??????? ??????? ???????????? ?????? . ? ???????? ????????????? ???????????? ?????? ????????? ? ???????????? ????????? , ? ?? — ? ??? ????? ?????? ??????????? ????????? . ????????? ????? ??? ??????????????? ? ?????? ? ????????? ? , ??????????????? . ?? ??????? ???????????? ????? ? ???????????? ???????? ????????????? ???????? — ???????????? ????????, ????? ?????????? ??????????? ; ????? ??? ? ?????????? ?????????. ????? ??????? ???????: ? ?????? ? ?? ???????? ??????? ????? — 300—350 ???, ? ????????????? ???????? — ????? 6 ???.
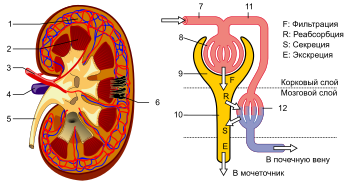
???????????? ????????? ?????? ?????? ???????????? ?? ?????? ???????? ??????? ??????, ?? ? ???????? ????????? , ??????? ??????? — ??????????? ????????????? ??????? ????????????? . ???????? ????????? ????????????? ? ????????????? ????? ??????????? ????? . ????????? ? ????? (? ??????, ? ? ?????????????? ????????) ???????????? ??????????? ???? , ???????????????? ???????????? ???????? ??????? H 2 CO 3 , ?????? ??????????? ???????????? ????? H + ; ??? ?????????? ??????? ??????, ??????? ? ???? ?????? ??????????? ?????? ??????? ??????????? ???????? .
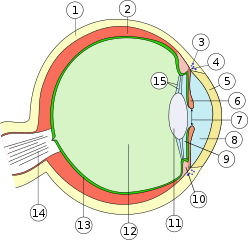
??????????? ??????? , ?????????? ???????????? ????????? ?????? ???? ?????????? ? ???????????? ?????????? ? ???????????? ?? ?? ????????? ???????, ????????? ? ????????????? ???????? ????????????. ??????????? ????? ?????????????? — ??????????????? ?????? , ????????? ?????????? ??????????? ???????? ?????????? ????? ????????????? ?? ??????????? ??????? . ??? ??????? ?? ??????? ? ?????? ?????????? , ? ????? ??????? ? ?????? ?????????? . ?????????? ???????? ??????????? ? ?????? ?? ????? ????? ? ?????????? ?? ? ?????????, ? ??, ??????????, ??????????? ????? ? ??????? . ?????? ?????? ?????????? ????? ????? ? ? ?????????????? ???????? — ?????? , ???????? ??????? — ??? ?????? ?????????, ?????????? ????????? . ?????? ????????? ? ?????????????? ????? — ???????? . ???????? ????? ?????? ( ??????? ) ????? ?????? ???????? ? ???????????? ????? ?????????????? ???????? . ? ???, ?????? ?????????????? , ????????? ? ?????????? , ??????? ??????, ?? ????????? ???????????, ?? ????????? ? ????????? ????????????? ????????? ? ?????????? ?? ? ?????????????? ???????. ??? ???????? ?????????? ??????? ?????? .
?????? ??????? ?????? ???????????? ????? ???? ?????????????? ( ???????? ): ?? ??????? ????????? ???????? ????? ????????? ?? ???????? ???????? ? ??????, ?????? ?? ???????? ????? ??????????? ?????????? ????? ??????? ? ????? ??????????. ????? ??????? ?????? ???????????? ?????? ???????? ????? ?????????????? : ?? ?????? ????????? ??????? ???????????? ???? ????? — ????? (???? ????????????? ?????????? ?? ?????????? ? ????? ?????? ????? ? ?? ???? — ? ????????????, ?? ?????? ), ????? ??????????? ??????? ???????????? ????? ?????????? ?? ????? ????. ????? ?????, ????? ?????? ????????? ???????? ? ??????????? ???????? ? ?????? ??????????? ? ???????? ??????, ????? ???????? ??????? ???????????? ? ?????? — ? ??????? ??????????, ? ??????? ??????? ???????? (?????????? ????? ?? ?????? ? ???????? ???????????) ? ?????? (?? ?????? ????? ????) ????? ???? .
????? ? ????????? ? ??? ????? — ?????? ?????????????? ????? ????????? ?????????????? . ????? ??????? ?? ?????? ? ????????? ????????? : ???????????, ?????????? ? ??????????? (??? ??? ????????? ???????? ?????????? ? ????????????? ? ??????? ?????). ?????????? (? ????????????? ???, ? ??????? ?? ?????? ???????????, ?????? ???????????? ????? ??????????? ???????? ????????? ) ?????? ???????????? ???? ? ???????? ??????????? ????? , ? ????? ????????? ? ??????????? ?????????? . ????????? ( ????? ???????? ?????? ) — ?????? ???????? ??????? ?????????. ?????????? ( ??????? ???????? ?????? ) ???????? ???????????????? ????? ?????????? , ?????????????? ??????? ????????? ?? ?????? ? ?????? ????????? .
??????????? ??????????? ????????????? — ?? ????? ??????, ?????????????? ????? (? ??????????? — ???????? ) ? ?????????? ???? . ?????????????? ????? ??????????? ??????????? ??????? ??????????? ??? ???????? ????????? ?????? ??????????, ? ????????? ?????????? ???? ? ?????????? ?????????? ?????? ???????????, ???????????? ????????. ? ??????????? ????? ??????????? ??????????? , ? ??? ??????????? ??? ????????????? ?? ???? ??????????? ??????? . ???? ?? ?????????? ?????????? ???????? ? ???? ????????? ??????? , ??? ?? ?? ???? ?????? ???????????? ??? ????????????? .
????????????? ??????? ????????? ??? ?????????? ? ??????????? ??????? ? ?????? ??????????? ? ?????? ??????? ????? ?????? ? ???????. ????? ?? ????????????? — ??????????? ????????, ???????????? ????????????? ????? ?????? ?????????? ??????? ????? ? ??????????? ?? ????????????? ??????? . ?????????? ?????????? ????? ???????? ????????? , ?????? ????? ????? ???????????? ?????????? ?? ????? ? ????? ? ?? ????? ? ????? . ? ????????????? ??????? ????????????? ??????????? « ????????????? ?????? » — ???????????? ??????? ???????, ????????? ? ??????????? ? ??????????????, ????????? ??? ??????????? ??? ????????????? ????????? ?????? ????? ???????? ????? ?? ??????? ?????????????? ???????????? ????????? ???? .
???????? ??????? ????????? ????????? ????????????? ????????? , ??????? , ?????? , ????????? , ?????????? ?????????????? , ? ????? ????????? ??? ?? ??????????? ???????????????? ?????? (????????, ?????????? ). ?????? ???????? ????? ??????????????? (??????????) ????????? , ??????? ??????????? ????????????? (?????????????) ??????????? . ? ???????????????, ? ????????????? ????????? ????? ????????? ??????? ??????, ?????? ??????????????? ???????? ??????? ????????? ??????? ????????????? .
??????????????? ???????? ????????? ???????? ??????????? ????? ????????, ??? ?????????????. ?? ????????? ? ???????? ?? ??????? ?? ???????? . ?? ????? ???????? ????????????? ???????????, ?????? ??? ??? ??????????????? ???????? ????????, ? ??????? ?? ?????????????, ?? ?????????? ?????????? ??????????? ?????? ?????? ??????, ??????????????? ?? ?????? ?? ??????????? ????????. ? ??????????????? ?????????? ????? ??????? ?????? ????? ????, ?????????????? ???????? ??????????????? , ??????????????? ?????????? , ??????? ??????????? ?????? ????? , ????????????? ?????????? ???????, ? ????? ??????????????? ?????????, ????????, ??????????? ? ???????????? . ?? ????????? ?????? ??????????????? ?????? ???????????? ??????????? , ??????? ????????? ? ?????????? , ? ????? ??????????? ??????? . ?????? ????????? ???????????? ??? ?? ?????????????, ??? ? ??????????????? ??????????. ?????? ??????????????? ???????? ?????????? ???????? ????? ?????????????? ??????? .
????????????? ???????? ????? ????????? ?? ?????? ?? ??????????? ???????? . ?? ???????? ? ???? ????????? ? ??????????? ?????????. ????????? ????????????? ???????? ????? ???????????? ????????? ????????????????????? ?????? ? ?????????? ??????, ??????? ????????? ??????????? ????????????? ????? ???????? ??????. ????? ??? ???????? T- ? B-????????? . ??? ????????? ??????? ???????????? ?????? ??????????? ??????? ?? ?????????? ??????? ?, ??? ??????????? ????, ???????????? ?????????? ?????????. ????? ????, ????????????? ???????? ????????? ???????? ????? ????????? , ? ??????? ??????????????? ?????????? ? ?????? ????????, ? ??? ????? ???????????? ? ???? ????????? ??? ???????????? ????? ??????? ? ??????????? ???????? ????? .
??? ?????? ????? ?????????? ?? ?????????????? ????????? ?????? ???????? ????? . ?????? ? ???????????? ?????? ?????????????? ???????? ????? ?? ??? ????????? ? ????? (?????????? ??????) ??? ???????? ? ??????? ????? ? ???????? ?????????? ? T- ? B-????????? ??????????????. ? ????? ? ???? ????? ? ??????? ???? ???????? ?????????? ??????????? ???????? . ? ??? ???????????? ????????? ???????????? ? ?????????????? . ? ???????? ?????????? T-????????? ???????????? ?? ??????????????????? ?????????????? ????????, ??????????? ???????? ? ???????????, ??? ???? ??????????? ??? ?????? T-??????, ?????? ???????? ??????? ?????? ? ???? ?????? . ?????? T- ? B-????????? ???????? ????? ?????? ?????????? ? ????????? ?? ????????? ?????????? ?????? : ????????? , ????????????? ???? , ? ????? ( ????. mucosa-associated lymphoid tissue, MALT ). ??? ???????????? ????? ????????? ?????????? ??????, ????????????? ??? ????????? ?????????-????????? ??????, ??????????? ?????, ????????????? ?????, ??????? ????? . ????? ????, ?? ????? ???? ???????????? ?????????? ????????????? ??????, ?????? ?????????????? ? ??????? ?????????, ????????, ????????? ??? ???????? ?????? .

????????????? ????????? ? ????????????? (?? ???? ????????????) ????????, ?????????????? ???????????? ??????? ??????????? ???? ? ???????? ?? ???? ?????????? ?????????-????????????? ????????? (?????????? ?????????? ?????????????? ?????? ????? ???????? ). ??? ???? ? ??? ?????? ??????? ?????????? ?????????????? : ????????????? ????????????? ????????? (???????? ?? ??? — ????????????? ???? ? ???????? ??????), ?????????????? ????????? ?????, ???????????? ??????????? — ??? ???????????? ???????? ????????? ?????? ?????????????? ????? .
?? ???????? ?????? ????? ????????????? ???????????? ???????? ??????, ? ? ?????? ???? — ????????? ???? ???? . ??????? ????? ????????? ??????????????. ???????????? ???? ? ?????????????? ?????? ???????? ??????? ?????? ??????????? ???????, ??????? ????????? ??????? ???????????? ??????-???????????? ????? . ???????? ???????????????? ?? ???? ????????? ?????????? ??????????? ?????????? ???????????????? ???????????? . ? ????? ???????? ???????, ? ?????????, ??????? , ???????? ????? ??????? ????? . ????? ????, ? ???????? ?????????? ?????????????? ??????? ????????? ????? ????????? ? ???????? . ???? ?? ????? ?????????? ? ???????? ?????????? ????????? ????, ?? ??????? ? ?????????????? ??????????? .
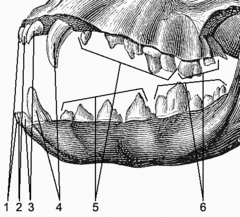
??? ??????????????? ??????? ?????????????, ? ??????? ?????? ??????????????? ????? (? 4 ????????: ????-????????? ???????? , ????????? , ???????? , ?????????? ) ? ??????????????? ?????? , ?????????? ????? ????????? ???????????????? ?????? ??? ???????? ??? ??????????????? ?? ????????? ? ??????? ???????? ??????????? , ??????? ??????????? ???????????? ???????? ??????????????? ????? . ??????? ????????????? ????????????? ???? ???????????? ???????????? ????? ??????????? ???????? ??????????????? ???????????, ??? ??????? ????????????? ( ????? - ?????????? , ???????? , ????? , ?????????? ) ????????? ? ???????? ??????????? ? ????????? ??????? ?????????? ???????????? ?????? .
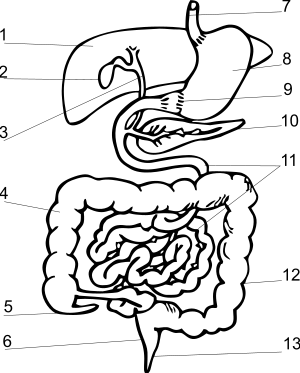
??????? ????????? ? ????????????? ?????????? ????????? ?????? ?? ??????? ?????????? ?????? ; ????? ????, ???????, ? ????????????? ???????????, ? ? ???????????? ???? ????? ??????? ? ?????? ??????????? . ???? ???? ? ??????????? ?????????????, ????? ??????????? ????????????? , ?????? ????? , ?????????? ? ?????????? . ??? ????? ? ??????? (?????????) ????????? ?????? ? ???????? ?????????????? : ???????????????? ?? ????? , ????? , ???????????? ? ???????? ???????? . ????? ?????, ?? ????? ? ??????? ? ?????? ????????????? ???????????. ???, ??? ???????? ?????????? ?????? ???????? ???????????? ???? ??????, ???????? ??? ???????? ??? ?????????? ??????????????, ? ?????????? ??????, ? ? ?????? ?????, ????????, ?????? ??????? ? ????????????? ??? ??????? ? ??????????? ?????? .
????? ????????? ? ??????? ??????? ???? ?????????????? ??????; ??? ???? ??? ??????????? ?????? , ??????????? ?? ???????? ?? ??????? ?????, ??? ????????? ?? ????????????? ? ??????????? ?? ????????. ??? ???????? ????? ??????? ???????? ( ??????? , ????? ), ???????????? ? ????, ??????????? ?? ???????? . ?? ??????? ??? ???? ???????? ? ?????? , ? ?????? — ????? ??????? ? ??????? . ????????? ? ??????????? ????????????? ???????? ??????? (????????????); ?????? ? ????????? ????????????? — ???????? ? ?????????? ??????? ? ?????? ??????????????? ????????????? ??????? — ???????? ??????? (?????????????) ???????, ????????? ?? ?????????? ???????????? ???????. ???????? ?????? ? ??????????????? ? ??????????????? ????? ?? ??????? ? ??????? , ?????????? ??????????????? ????????: ? ?????? ???? ??????? ???????? ???? ???????????? ???????? ? ???????? ??????????????, ? ??????????????? ?????? ???? ???? ? ??????? ?????????? ??????, ??? ???? ? ???????????? ??????????? ??????????? ???? .
???????? ?????????????? ?????????????? ?? ?????? ? ??????? (? ?????????? ????????? ?????? , ????????? ? ?????? ????? ) . ???????? ????? ???? ?????????????? ? ?????? ?????????, ????? ?????? ???????? ???????? ??????????? ???????? ??????????? ? ????? . ??????? ?????????????? ???? ????????? ? ??????? ????????, ??? ??? ???????????? ?????????? ????????? ? ???????? ?????????????? — ????????, ??????? ? ??????????. ??? ?????, ?????????? ?????? ???????????? ?????, ????? ?????? ???? ? ???? ????????? ?????? ?????? ?????, ????? ???????? ??? ?????? ????????????? ? ?????????? ?????; ?????? ?????? ????? ??????????? ????????????? ???????? ??????? ?????????? ????????????? ?????, ??-?? ???? ??? ?????? ??? ? ?????? ???? ? ???????? ??????? . ? ????????? ????? ??????? ????? ??????????????, ???????????? ? ?????? ????? ? ????? ????????? ?? ????????? ????? ???????? ????????? .
????? ????????????? , ??? ? ? ?????? ???????? ???????????, ??????? ( ??????????????? ) . ??? ?????? ????? ?????????? ????? ? ????????????? ? ?????????? ???????, ?? ????? ????????????. ? ???????? ???????? ???? ?????? ????? ????????????? ??????? , ? ???????? ????????? ??????? ?????????? ?????????? ?????? ????? ? ?????????? ???? , ????????? ????? ?? ???????????? ? ??????? ?????? . ?? ???? ???? ?? ??????????????????? ?????? ??????? ?????? .
????? ?????????? ?????????? ???? ? ????????? ? ??????? ?? ???? ????????? ???????? ?????? ???????. ????????? ?????????? ????????? ?????? ???????? ???????? ? ??????? ??????? . ???????? ?????????? ? ?????? ?? ??????? , ? ??????? ??????? — ??????? ??????? ??????? . ??? ???? ? ????????????? ???????? ?? ?????????? ? ???? ????? ??????????? ??? ??????? ???????? (? ???????? ? ???? ??????????? ????????). ?????? ?????, ???????????? ???? ? ????????? ?????? ??????? ?????? ? ?????? , ? ????? ?????? .
????? ????????????? ????? ???????? ???????? ???????, ??????????? ? ????????????? . ??? ????? ???????? ?????????????? ???? ??? ??????? ???? ? ??????????? ????????????????? — ??? ?? ???????? .
??????????? ??????? ?????????? ?????? ?????????? ???????? , ???????????? ?????? ????????? ??????? ????. ??? ?????? ?? ????? ???????? ???????? ? ???????? ?????????????? ??? ??????? ? ????? (???? — ? ?????) . ? ????????????? ??????????? ??????? ????????????? ?????? ? ????????? ????????? ?????? ???????????, ????????? ?? ??????? ???????? ??? ?????? ????????, ??????? ????????? ?? ?????????? ? ????????????? .
? ????? ????????? ??????????? ????? ??????? :
????????????? ?????? ? ??????? ??????, ????? ??????????? ????????, ???????????? ? ??????????? ????????: ?????? ????????, ????????????? ?????? ???????? ????? ??????????????? ???????? , ? ??????? ?????? ??????????? ?????? . ? ????? ? ???? ????????????? ?????? ? ??????? ?????? ??????? ? ??????? ????????? ???????? . ????????? ???????? ?????????? ? ????????????? ? ?????????-???????? ??????, ??????, ?????? ? ?????? ???????, ?? ??????????? ? ??????????? ??????? .
?????????????? ? ?????????????, ??? ? ? ?????? ?????? , ??????????. ??????? ?????? ????? ????????????? — ??????? — ?????? ????? ? ??????? ????, ? ??????? ?????? ????????? ??????????? ? ???????? . ?? ????? ???????? ????????? ?????????? ????? ??????? ???? ???????? ? ?????????? ????? — ??????? ??????? ????? ???????? . ???????? ???????? ????? ????????????? ?????????? ?????????? ( ??????????? ???? ); ?????? ? ??? ??????????? ?????????? ???????????? ?????????? ????? , ????????????? ? ???? ??????? ????? ( ?????????? ???? ? ??????????? ????????????? ? ????????????? ???? ? ???????? ? ????????? ????????) .
??????? ?????? ?????? — ????????? — ????? ??????????? ???????? ?????; ? ???? ????? ??? ? ??????? ???? ????? ????????? ???????? ? ??????? ????, ?? ? ??????????? ????????????? ?? ???????? ?????????? ? ????????????? ??????? ???????? ??????? — ??????? . ?????????????? ??????????? ????????????? , ?????????? ? ???????? ????????? , ???????? ?????? , ??????? ?? ????? ???????? ???? ????? ????? ????? ???????? ?? ????????? ????? (? ????????????? — ? ?? ?????? ); ????? ????? ? ??????? ?????? ???????? — ??????????? ????? — ? ?????????? ?????????????? ; ?????????????? ?????? ??? ?? ???????? ????????? ? ????????????? ? ????? .
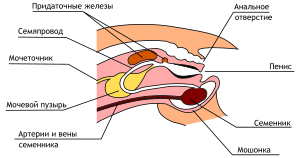
??????????? ????? ? ????????????? ?????????? ? ????? — ?????? ?????? ???????? . ??? ???? ? ???????? ? ???????????? ?? ????? ???????????? ????????? ???????? — ????????????? ?????, ???????????? ????? ????????? ???????? ???????? ???????? ( ??????? ) ? ????????????? ???????? ????? ? ?????????????? ????????? ???????? ?????????? ? ???????????? ??????????, ? ????? ????????? ??????????? ? ????????? ??????. ?????? ? ???????? ???????? ?????????? ?????????? (??-?? ???? ? ??? ?????? ?????????????? ????? ???????, ? ??????? ????????? ?????? ????????????); ? ???????????? ?? ??? ??????? ????? ??????????, ??? ????????? ??????????? ???????? ?????? ??????????????? ????????, ? ??????? ???????? ???? ??????? ??????? ?? ??????????????? ??????? ??????? ????? .
????????????? ????????????? ???????????? ???????????? ?????????????? ? ???????? ? ????????? ????????? ?????? ? ? ????????? ?????? ?????????. ?? ???? ????? ??????????? ?? ??? ? ???????? ????? ???????? ?????? ???????????? ????, ??? ?????????????, ?????? ??????? ????, ???????? ? ???? ? ??? ?? ?????, ????????? ? ????????? ?????????? ????? — ???????? ????????????? ???????; ? ?????? ???????, ????????????? ?????? ???????, ??????? ? ??????? ???????? ? ??????? ?? ?????? ?????? ?????, ????? ????? ?????? ??????? ??????? ??? ( ????????????? ???????? ) .
? ???????????? ? ?????????????????? ? ????? ? ???????????? ????? ????? ????????????? ???????? ????????? ?????????? ????: ????????, ?????????, ??????, ????????? ? ???????? ????? (? ????? ?????????? ??????) .
???????? ????? — ???????? ???????? ?????? ?????????????. ? ???????? ?????? ?????? ????? ???????? ??? ???????? ?????: ?????? ?????? ? ?????? ???????? ????????????? . ? ??? ???, ? ??? ?????? ???????????? ????????? ???????????? ?? ???????? ????????? ???????? ?????? ? ??? (? ?????????????? ?? ?? ????????????? ??????? ) .
??? ?????? ?????????????? ?????? ?????????????. ????? ??? ??????? ? ????, ? ??????? ?? ?????? ? ????????. ????????? ?? ?????? ?????? ????? ?????? ???????? ????? ????? ( ???? , ?????? , ????????? ????? , ???????? , ??????? ); ?????? — ??? ?????????? ????????????? ????? — ???????? ? ? ??????? ?? ????????, ??? ??? ????? ???????? ???? ? ???????? ( ?????? , ????????? ??????? , ???? , ????????? ) .

?? ????? ?????????????? ?????? ???????? ????????? ???????? ??????????? ( ????? , ?????? , ??????? ). ????? ?? ??? ?????????????? ? ???????? ???? ?? ??????? ???????????. ??? — ??????????? ???????? ( ?????? , ????? , ???????? , ?????? , ??????? , ?????? ), ? ????? ????????? ?????? ( ???? , ??????? , ?????? ). ?????? ?????? ??????? ??????????????? ??? ???????? ?? ??????? ?????? ??????????? («?????????????? ???»): ???????? ?????????? , ?????? ??????? , ?????????? , ?????????? ??????? , ???????????? , ?????????? , ????????????? ???? . ???????, ?????? ?????? ???????? ?????? ??? ??????? ???????? ????? ? ????????? ?????? ? ??????????? ?????, ??????? ????? ? ?????? ?????, ???????????? ???????????? ( ????? , ??????? , ?????? , ????????? ?????? ). ?????? ??? ????????, ? ?? ????????? ???????? ? ????? .
??? ????????? ????? ???????? ?????? ( ???????????? ????????? , ???? , ????? ?????? , ?????? ) ?????????? ??????? ????? , ???????????? ????? ????????????? ????????? (????- ). ? ???? ?????? ???? ?????????? ?????????????? ? ????? ? ?????-???? ???????????? ????? ???????????, ? ?????? ???? ????????????? ???????????? ????????????? ???????? . ???, ??????, ????? ???????? ?????????? ?? ???????? ????? ?? ??????? ? ??????????? , ?? ?????? ?????? ? ??????? ?? ??? ??????????? ? ?????? , ? ?? ??? ? ?????? ?? ??????? .
?????????? ???????? ???? ????? ???????? ?????? ????????? ?????? — ?? 105—115 ??/?. ???????????? ???????? ???? ??????????: ? ???? ? ?????? ??????? — 75—80, ?????? — 65—70, ????? , ??????? ? ?????? — 60—65, ???????? ? ????? — 55—60, ?????? ???? — 50—55, ?????? , ?????-?????? ? ??????????? ??????? — 45—50, ?????? — 40—45, ?????????? ???????? — 35—40 ??/? .

?????? ????????????? ?????????? ???????? ?? ??????????? ? ?????? ????? ????????????? ????????? ????? ????????? ? ?????????? ???????. ? ????? ????????????? ?????? ????????? ?????, ?????? ????? ??? ????? ??????? ????, ?? ?????????? ???? ?? ??????????? ?????: ??????? , ??????? , ????? , ??????? , ???????????? , ?????? , ??????? , ?????????? , ??????? . ?????????????? ? ????? ??? ?????? ? ??? ???????? ?????. ?????? ??? ??????????? ? ??????? ??????????? ????? ????????? ? ???????? ??????????? ??????????? .
????????? ????????? ????? — ??? ????????? ???????????????????????? ?????? ?????????????, ??????? ???????? ??? ??? ???????? ????? ????? ? ????? ?????, ?????? ??? ???? ????: ???? ????????? ? ????? ??????????????, ???? ????????? ????? ????????. ????????????? ?????? ?????? ??????????? ? ?????? ???????. ???????? ????? , ?????????? , ????? , ?????? ???????????? ????????? ???? ???????? ????????? ???????????? ? ???????? «????????????» ???????, ? ??????? ? ?????????? ?? ?????? ?????, ? «?????????» ?? ???????????? ?????? ??????? . ??????? ?? ?? ????????? ???????????? ????????? ???????????? ? ????????? ??????, ? ????????? ??????? .
? ????? ????? ??????????????? ???????????? ????????? ?????? ????????? ?????????????? ????? ???? ? ??????????? ??????? ? ?????? ??? ????????? ???????? ???? ? ????? ??????? .

????? ????????????? ??????? ?????? ?????????? ?????????? ????? — ????, ????? ??????? ???????? ???????? ?? ????, ???????? ? ????. ????????? ?? ????? ???????? ( ????? , ????? ??????? , ??????? ??????? , ???????? ?????????? ) ????????? ??????? ? ??????, ?? ????????? ????? ???????? ?????. ?? ????? ? ?????? ?????? ??????????? ? ???, ??? ???? ???? ??? ???????? ?? ?????? ?? ????, ?? ? ? ???? . ????????????? ???? ?????????? ???? ????????????? ?????? ????????? ??? ? ???????? ??????? : ????????????? ? ??????????? ?????? ????? ??????????, ? ?????????, ??? ????????? ???????????? ?? ??????? ??? .

??????????????? ????????? ??? ????????? (?? ??????? ????????? ? ????? ? ????) ?????? ?????????? ?????? ?????? ??????? ????? ??? ????? ???????? ???????????? ????????? ????? ???????? ???????????. ????? ????????? ??????? ? ???????? , ???????? ???????? , ???????? , ?????? ???????? ( ?????? , ???????? , ???? , ???????? ????? , ??????? ? ??.), ????? , ??????????? . ? ?????? ? ??????? ????????? ???????????? ????????? ???????? ???????????? ???????? ?? ??????? ????????. ??? ????????? ????? ?????????? ????? ?????????? ??? ? ????? ???????, ???????? ?????????????? ??? ????????? ???? ? ???????, ????????? ??????, ??????? ??????? ????, ??????????????? ?????????, ??? ???????? ?????????? ???????? ???? .
??? ????? ??????? ??????? ?????????????? ? ????? ? ?????? ????? ????????????? ?????? ? ?????????? . ??????? ???????? ????????? ????? ???????? ? ??? ??????? ?????????, ??? ?? ?????????? (? ?????? — ?????? ??????) ???????????? ? ????? . ???? ??? ???????? ???????? ?????? ? ????, ?????? ??????????? ? ???????????? (? ?????? ? ????????) ??? ??? ?????? ????? .
????????? ??????? ??????? , ??????? ?? ??????????? ?????? ?????, ???????? ?????? ? ???????????? . ? ??? ????????? ??????????? ??????? ??????, ???????? ?????????? ???????????? ? ????????, ? ?????? ????????. ???????? ???????????? ??????? ?????? ???????? ????????? ??????? . ???? ? ???????? ? ?????? ???????????? ????????????????? ?????????? ??? ????? ????? 8—10 ???., ? ??????? ? ???????? — 12 ???., ????? — 15 ???., ????? ? ????????? — 16 ???., ?? ? ????????? — 20 ???., ???????? — 75 ???., ? ? ??????????? ??? ????????? 2 ?????. ??????????????????? ?????? ?????????? ???????? ?? ??????? 988 ? , ? ????????? ????????? ?????????, ??? ?? ???????? ??????????? ???? ?? 3 ?? .
???? ???????, ??? ?????????????? ???????????? ??? ????????? ?????? ???? ????????? ? ???? ??????? ?????????????? ??????? ???????????. ?????????????? ???????????? ? ?????? ??????? ?????? ????? ??? ??????????. ??????? ?? ??????? ?????? ????????? ???????????? ?? ??????? ??????????? ? ???????? ????? ??????? ? ????, ??????? ?? ?????, ?? ??? ?????? ??????? ?? ???? .

? ????????? ?????? , ????? ????????? ?????????????? , ??????? ?????????????, ?????????? ???????? ????? ????? ????? ? ?????? ????????, ??? ??? ???????? ????, ????????? ?? ??????, ?????????? ?????? ??? ??????????? ? ??????. ???????? ??????? ???????????? ? ??? — ??????? ?? ??????? ? ?????? ???????? ? ?????? ? ????? ?? ????? .
?????????????? ??? ????? ?? ???????? ? ????????????? ????????????. ?????? ?? ??? ?????????? ??? ??????? ??????? ???????????? ?????; ??? ???? ???? ?? ??? ????? ??????? ???????? ?????, ??????? ?????????? ??? ?????????? ? ????????? ??? ??????? ( ???????? ?????? , ???????????? ??????? , ????????? ??????? , ????? , ???? , ????????? ?????? ) , ? ?????? ????? ?????? ???????????? ???????, ??????????? ????????????? ????????? ???? ?? ????? ( ??????-?????? , ??????? , ?????????? ???????? , ?????????? ???? , ????????? ????????? , ??????? ) , ? ??????? ????? ???????????? ? ? ???????????? ?? ????????? ( ????? , ??????? ) .
??? ????????? ?????? ?????????? ??? ??????? ???????????? ?????????? ? ?????? ????????? ????????, ?????? ???? (? ??????? , ????????? , ?????? ??????? ) ??? ??? ?????? (? ????? , ???????? ) ????????????????? ????????? (?????????, ??? ???????? ????? ?? ?????? ??????? ???????????????? ?????????, ??? ????????? ???????????? ??????? ???????????). ??? ???????, ????? ???????????? ????????? ?? ????? ????? ???????? ????????????. ?????? ??????? ? ????? ?????????? ??? ???????????? ?? ?????? ? ?????????????? ? ?????? ?? ?????? ???? ???????? ??????????, ? ????? — ????? ? ???????????? ????? .
???? ????? ? ???? ?????? ????????? ??????????? ????????????? ?? ????? ? ?????? ??????, ?? ????? ?????????????????????? ???????????? ????????????? ?? ????? ?????, ? ??????? ????? ??????????? ???????? ????? ????????? .

??? ????? ????? ???????? ????????????, ????????? ??????????????, ???????? ????? : ??? ????????? (???????????), ??? ? ???????? (? ??????? ???????). ?? ??????????? ? ??? ??? ???? ?? ???? ?????????????? ??????, ?????????? ????????????? ????????? ??????? ???????? ????????? ?????, ????? ???????? ?????? ???????? ?????????????? ????????????? ? ????????? ????????? ????? .
????????????? ????? ????????????? ????????? ? ?????? ?????????? ? ???????, ?? ???????? ????? ?????? ????????. ???? ???? ? ??????? ??????????? ????????? — ???????? ???????, ??????? ??????? ?? ????? ???????? ????? ????????? ? ??????? ?????? ? ????????? ?? ?????? ??????????? ??? ?????? ?? ??????? ?????, ?? ? ????????? ???????? ? ???????. ? ????? ?????? ???????????: ??????????? ????????????? ?? ??????? ??? ?????, ????????? ??????????? ???????? ( , ????????? ??????? , ?????????? ??????? ?????? , ?????????? ??????? ?????? ), ????????? ??????? ( ?????? , ??????????? ??????????? ), ??????????? (? ????????? ??????????? ????????? ????????? ??????????? ????????, ????????? ? ?????). ???? ? ?????????? ?????? ????????? ?????? ????????? 60 ?, ?? ? ??????????? — 140 ? .
?
????????? ????????? ??????
??????????? ?????????????? ????????????? ??????
??????????
, ? ??????? ???????? ?????????? ?????????? ? ??????? ???????? ??????. ?????????? ???????? ?????????? ?????, ?????? ? ? ??????? ?????????????? ????????????? ? ???????, ???????? ??? ?? ???? ???? ? ???? ???????????
.
???????
?????????? ?????
????
, ??? ? ? ????, ???????? ??? ???????????? ???????? ??????? ????
. ???????? ????????? ???????????? ????????????? ???????? ????????????? ?????????
??????????????
?, ? ?????????,
??????????? ????????????
: ??? ????? — ?????? ? ?????????????, ?????? ??????? ???????? ?????? ?????????? 40 ??/?, ? ???????????? — 100—105 ??/?
.
?? ???? ????????????? ???? ????????????? ????? ???????????? ?? ??? ???????? ??????: ???????????????? ? ?????????? . ?????????? ??????? ??????? ??????? ? ???, ??? ???? ???????? ???? ????????????? ???????? ????????????? ?????????? ??? ????????????? ?????????; ? ??????????? ? ??????? ???????????? ? ????????????, ? ???????? ?????, ??? ??? ???? ???? ???? ? ???????????? ??? ??? ?????? . ???? ????? ???????????? ?? ????????, ??????? ? ???????? ???????? — ?????? ????? ??????? , ???? , ?????? , ?????? , ????? , ????? ????? ? ??.; ? ?? ??????? ????? ??? ????? ???????????? ???? ?????? ?????????????? ? ??????????? ????????, ?????, ??????, ?????, ??????. ? ?????? ???????, ????????? ???? ???????????????? ?? ???????????? ?????? ????????????? ?????? ?????? .
????? ???????????????? ????????????? ?????????? :

????? ?????????? ????????????? ?????????? :
? ????????????? ????????????? ? ??????? ????? ?????? ????? ???????? ?????????-??????????????? ????????????, ??????????? ??? ????????? ????? ????????????? (????????? ??????? ?????? ????? , ????????? ????? ? ??????? ??????? ???? ? ?????????? ?????????? ??????????? , ?????????? ? ?????????? , ??????????? ????????????? ?????????? ????????????, ????????? ?????????????? ????????, ??????? ?????????, ????? ?????? ???????? ? ???? ?????? , ??????? ?????? ????? ???????? ? ??.). ??? ??????????? ????? ??????????? ? ??????? ????????? ?????????, ??? ?? ???????? ? ??????? ??????? ?????????, ??????? ?? ??? ???? ????????? .

????? ????????????? ?????? ????????????? ??????? ????????? , ?? ???? ???????, ??? ??????? ?????????? ????? ?????? ???? ?????????? ???? ?? ?????. ? ????????????? ???????? ??????? ???????? ?????????? ?? ????????? ?????????:
????? ??????????? ?????????????? ? ????????????? ????? ?????? ?????????. ???????? ????? ???????? ?????????? ??? ????????? ????? (? ??????? ???? ????????? ????????????????? ????? ? ???????, ???): ????? ??????? — 1 ???. (1—1,5), ??????? ????????? — 1,5 ???. (1—1,5), ??????? ???? — 2—3 ???. (????? ????), ????????? — 3 ???. (4—7), ???? — 1 ??? (7—9), ?????? — 1—2 ???? (10—12), ???? ? ???? — 2—3 ???? (15—20), ????? ??????? — 3—4 ???? (30—50), ????? — 4 ???? (12—13), ????? ??? — 4—5 ??? (?? 20 ???), ????????? ???? — 10—15 ??? (70—80) .

?????????? ????????????? ?????? ?????????? ? ????????????? ??????? ???? . ? ????????? ? ??????? ??????? ???????? ? ??????????? ???????? ??????? ?????? ????????? ????? ????????? ???, ? ?????? ??????? — ????????? ????????????? ???????, ?????????? ???????? ???????? ???????? ? ?. ?. ??? ?????? ????? ???????????????? ???? ? ????? ????? ????????????? ???????? ?????????? ????????? ?????, ? ?????? — ?????. ?????? ????? ?????????? ???? ????? ?? ??????, ?????? ? ??????? ?????? ??????????? ???????????? ??????? ?????. ?????????????? ? ???????? ???????: ? ????? ? ???????? ????? ?????? ? ?????, ? ?????, ? ?????? ????? ?????? ?????, ? ? ????? ?????? ? ????? ?????? ?????????? ??????????? ?????? ???????? ????? .
??? ??????????? ????? ????????????? ?????????? ????????? ? ????? ????????? . ? ???????? ????? ????? ( ????????????? , ??????? ???? , ?????? ??????? ? ?????? ) ????? ????? ?????????? ?? ?????????? ????? ????? ????. ? ??????? ??????? ? ??????? ???????? ?? ????? ??????????? ?????????? ?????? ??? ?????? ? ???????????? ???????? ?????? ?? ?????, ? ? ????????? ?????? ? ???????? ????????? ??????? ????? (???????? ????????, ?????? ?????) ?? ?????????? ?????? ? ????? ? ???????? ?????? ? ??????????? ????????????. ?????? ??????? — ?????????? (?????????????), ????????? ? ???? ??????? ????? ? ????????? ??????? (????????, ? ??????????? ?????????? ) . ??????????? ???????? ???????? ?????? ? ????? ?????????? : ??? ??????? ?? ?????????? ???????? ?????? ? ???????????? ?? ???? ?????????? ?????: ?? ????? ????? ?????????????? ?????, ??????? ? 1,5—2 ???? ??????????? ?? ???????? ????????? ?????? ????? ? ????? ??? ????? ???????? ? ????????? ?????? ? ???????? ? 2—3 ????????????? ???????, ??????????? ?? ?????????????; ????????? ???????? ????? ????? ?????? ?????????? ???? ? ?????? ? ????????????? ??? .
??????????? ??????? ?????? ???????? ?????????? ????, ?????? ???? ?????? ?????????? ?? ???????? ???? ( ?????? , ?????? , ????? , ????? , ??????? ), ???? — ?? ????????? ??? ( ????? ). ?????? ???????? ???? ??????? ????? ???????????? ???? ?? ??? ????? .
?? ???? ??????????? ????? ????? ?????????? ??????????? ? ???? ??????? ???????? ?????. ? ?????? ????? ??? ?????????????? ?????? ??? ?????? ??? ???????? ?????????. ? ??????????? ????????????? ????? ?????? ? ??????? ?????????; ????? ?????? ?????? ????, ? ????? ?????? ? ?????? ????????????? — ????. ??????????? ???????????? ?????? ??? ?????, ?? ????? ????????? ????? ( ???????????? ??? , ?????? ? ??.) ????????? ??? ????? ???????????? ?????????, ???????? ?? ???? ????????? ????? ???? ???, ??? ?? ??????? ?????????? ?? ???? ? ???????. ????? ?????????? ?????? ? ??????? ?????????: ????? — ???? ? ?????????????? ????????? (?????? ??????), ???? ? ???????????? (??????? ?????), ? ??????????? ? ??????? — ???? ???? ???????; ??????????? ??????? ???????? ? ?????????? ? ????? ???????????? ????????? ??? ?? ? ??????? ????? ?????? ? ?????????? ????????. ? ????????? ??????? ????? ??? ????? ????? ?? ?????, ??????? ?? ??? ????????? ??????, ? ???????, ???????? ??????? ?? ??????, ??? ?????????? ?? ????? ?????? .
?? ??????? ???????????? ???? ????????????? ????? ?? ????????????? , ??????? ?????? ????????? ??? ? ??? (??????????? ???????? , ????????????? , ?????? , ???????? , ??????? ????? ) ??? ??? ? 2—3 ???? ( ????? ??????? , ????? , ????? , ????? , ????? , ??????????? ?????? ????? ) , ? ????????????? , ???????? ????????? ??? ? ???? ( ?????-?????? ? ????? ???????? ?? 2—3 ??????, ??????? — ?? 3—4, ??????? ???? — ?? 6—8) . ???????? ? ???????? ???????: ???? ? ??????, ?????, ??????, ??????, ?????? , ??????????? ??????? ????????? ?? ?????? ????????, ?? ? ?????? ? ???? , ?????? , ?????? ??????? ?????? ?? 2—3 ????????, ?????, ?????, ????? — ?? 3 ?? 8, ??????????? ???????? — ?? 8 ?? 18 . ???????????? ??? ???????????? ????? ????????? ? ????? ?????? ?????? ? ????????????? ??????? : ? ????? ????? ?????????? 32 ???????? , ? ?????? ? ?????? ?????? ? ????????? 31 ???????? (??? ???? ????? ?????? ? ????? ??????? ????? ???????? ?? 29 — ??????, ??? ? ?????? ??????? ??????????????) .
??? ?????? ????? ?????????? ??????????? ???????? ??????? ?? ?????????????? ????????. ???, ? ?????????????? ??????? ??????? ???????? ??????? ?????????? ? ??????????? ? ??? ?? ?????, ?????????: ? ???????? ???????? — 4,0; ? ??????????? — 6,9; ? ??????????? ?????? — 8,2. ? ???????? ??????? , ?????????? ?? ???-?????? ??? , ??? ?????????? ? ??????????? ??????: ?? ?????? 1,0—1,5 ???. ? ??? ????? 4,6; ?? ?????? 1,7—2,0 ???. ? — 4,4; ?? ?????? 2,5—3,0 ???. ? — 5,4; ?? ?????? 3,2 ???. ? — 5,6 .

??????? ?????? ????? ? ?????? ???????????? ????????????? ? ???????? ? ? ??????? ????? ????????? ????????????? ????????. ? ?????? ??? ???????? ?? ??????? ????? ?????????? ???????? — ?????? ? ?????? ???????? ? ??????????, ?? ????????? ? ???????, ??????????? ?????? ? ?????????. ????? ??? ?????????? ??????? , ?????????? ?????? ???????? ? ????????????? ?????? ? ??????? ????, ? ? ?????? ????? ????????????? ??????????? ???? ??????????? ??????????? ? ????????? ?????????? ??????. ? ?????????? ???? ??????? ????????? ?????? ??? ????????? ?????, ?????????????? ????????? ?????? ???????? ? ???????????? :
| ?????? ?????? ????????????? | |||||||
|---|---|---|---|---|---|---|---|
| ???? ???????? | ?????? ??????, % | ?????????????? ???????? | |||||
| ???? | ????? | ???? | ????? |
???????????
???????? |
???? / 100 ? | ??? / 100 ? | |
| ?????????? ??????? | 88,0 | 4,6 | 3,4 | 6,7 | 5,3 | 76 | 318 |
| ????????? ???? | 78,1 | 4,9 | 11,6 | 4,7 | 0,7 | 143 | 599 |
| ?????? | 67,2 | 13,9 | 15,3 | 2,1 | 1,5 | 202 | 846 |
| ??????? | 87,1 | 0,9 | 4,5 | 7,1 | 0,4 | 72 | 301 |
| ????? | 82,2 | 9,1 | 3,3 | 4,9 | 0,5 | 86 | 360 |
| ?????? | 76,4 | 7,4 | 10,7 | 3,3 | 2,2 | 139 | 582 |
| ??????? ????? | 34,6 | 8,9 | 53,3 | 0,1 | 3,1 | 516 | 2160 |
| ?????? | 88,8 | 2,5 | 1,9 | 6,2 | 0,6 | 52 | 218 |
| ?????? | 81,2 | 4,8 | 6,8 | 5,5 | 1,7 | 102 | 427 |
| ??????? | 58,3 | 6,8 | 33,0 | 1,1 | 0,8 | 329 | 1377 |
| ???????? ????? | 67,7 | 10,1 | 18,0 | 2,8 | 1,4 | 214 | 896 |
| ???? | 82,0 | 4,6 | 7,2 | 4,8 | 1,4 | 102 | 427 |
| ???? | 86,7 | 3,2 | 4,5 | 4,3 | 1,3 | 70 | 293 |
| ?????? | 87,3 | 3,2 | 3,9 | 4,6 | 1,0 | 66 | 276 |

? ???????????????? ?????????? ?????? ?????? ????????? ??????????? ? ? ????????? ?????????????? ?????? ? ???? ?? ???? (? ??????? ????????? ??????????? ??????). ??? ???? ???????? ?????? ?????????????? ???????? , ??? ????????????, ?????? ? ??????? ????????????? — ?????????? ? ????????????; ????? ???????? ??????? ???????? ??? ????????? ? ????????? ????? .
???????????? ????????????? ??????? ? ?????? ????? ????????????? ????????? ? ??????? ????????. ??????? ??? ????? 7—8 ???? ???????? ???? ?????, ???? ?????? ??????????? ?????? ??????????. ? ???????? ? ?????? ???????? ?????? ???????????? ???????, ?? ? ????????? ????? ??? ???????????? 4—5 ???., ?????? ? ?????? — 9—10 ???., ????? — ?? ????? ???? .
????? ???????? ????????? ?? ??????????????? ??????? ??? ???? ??????? ???????????. ????? ????? ?????? ??????? ?? ?????? ????? ?????????????? ? ??????? ???????. ??????? ???????, ??????????? ??? ????????????? ???????? ? ???????????? ????????, ??????? ???????? ???????? ?????????. ???, ????????????? ???????? ????? ??? ? ?????? ???? ? ????????? ????????? ?? ???????, ? ????????????? ??????????? ????? ?????? ??? ?? ????? ?????????? ?????????? ????????. ?????? ?????? ?????? ???????? ? ?????????? ? ?????? ???????? , ? ??? ???????? ????????? ????????? ?????????? ???????? .
?????? ? ???????????? ????????? ????????????? ? ???????? ????? ?? ??????, ???????? ??????????? ??????????, ???? ? ???? ?????, ???????? ??????????, ???????????? ? ?????. ????? ???????????? ????? ????????? ???????, ????????? ?????? ?????? ????, ??????????????? ?????? ? ????, ????????? ?? ????? ????? ? ?????? ?????????????, ???????? ?? ??????, ?????????? ?? ???????????? . ???????? ?????????? ??? ????????????? ??????????????? ???????? ????????? ? ????????? ????????? ???????????? ?????????? ??????????????? ????? .
? ?????????????, ??????? ?? ??????????? ? ?????????? ?????? ?????? ???? , ????? ??????? ??????? ???? ????? ? ??????????????? ? ???????? ?????????? ????????? ??????????. ?????? ?????? ???????????? ??????????? ???????????? ? ???????? ?????????????? ??????? (??? ?????? ???????? ? ????????? ????? ??? — ????? ????? ? ?????? ????). ? ???????? ???????? ?????? ???? ????????? (? ?????? ???????????? ????????????) ? ?????? ??????????, ??????????? ??? ???? ??? ????? ???? .
????????? ????????? ?????? ???????????? ????? ?????, ???????? ???????? ??? ???????????? ? ?????????? ?????????. ? ??? ????? ???? ????????? ??????????? ???? ????????? ? ?????????, ?? ????? ????? ?????????? ????? ????????. ????? ??????????? ????????? ?????, ?? ??????????? ? ??????????? ??????, ????? ???????????? — ? ?????, ???????? ??????? ?????? .
?????????????? ????????????? ? ??????????? ???? (?????? — ?????? ??????????????? ???????? ????) ????????????. ?????? ????? ?????? ??????????? ??????? ??????? . ???, ????????? ? ?????????? ??? ? ???????? ????? ? ???? ?????????? 10—15 ?, ? ??????? — 750—800 ?). ??????? ????? ???????????? ?????? , ??? ??????? ????? ?????? ?????? ??? ????????? ????? ??????, ? ? ?????? ??????????????? ????? ????? — ?????-?????? , ?????? ????? , ????????? ? ??. — ?????? ??? ?????, ??? ?????? ??????? ???????????? ?? ????? . ????? ????, ????????? ????????????? ????????? ???????????? ???????? ???????? ?? ?? ( ???????? ????? , ?????-?????? , ????? ? ??.), ?????? — ???????? ???? ( ?????? ???? , ??????? , ?????? , ??????? , ????? , ????? , ?????? ? ??.) . ?????? ???????? , ?????? , ??????? ???????????? ? ???? ? ????? ??????? ?????? (?????, ????) .
?????? ????????????? ??????? ?? ???? ? ?????? , ??????????? ??????? ??????? ??????? ? ??????, ????? ????? ???? ?????????. ????????? ?????? ??? (?????????????? ??????), ??? ??????? ?????? ???????????, ??????????? ???? ? ??????????? ?????????? ??????? ????????????? ( ??????? , ??????????? ?????? , ????? , ??????? ? ???????? ?????? ????? ???????), ? ????????? ?????? ? ??????? ??????????? ? ???????????? ????????? ??????????? ???? ? ????????????? ??????????? ( ??? , ??? ??????? ????? , ????? , ??????? , ?????? ). ? ????????? ???????? ???????? ? ?????? ??????, ???????????? ? ??????????? ??????? .

??????????? ??? ????????????? ??????? ??????? ???????? ?????? ??????? ???????????? ????????????? ??????? ????? ?? ?????????. ??-??????, ???????? ?? ????????? ? ??????? ???????????? ????????? ? ??????????? ? ????????????? ???????? ????????????? ????????????, ????????? ??????? ???????? ?? ??????? ??????????? ????????? ? ?? ??????? ?????????? ( ?????????? ). ??-??????, ?????????????, ???????? ?????? ???????????? ? ???????? ?? ??????? ???????? ???????? ???? ????????? ????????? ?????, ?????????? ?????????? ???????????? ? ??????? ??????? ? ?????????? ??????? ???????????, ? ??????????? ???? ???????, ? ????????? ???????? ????????? ? ?????????? ??????????????? ????? .
?????????? ??????????? ????????? ????????????? ??????????? ? ????????? ?????? ?? ?????. ?????? ?? ??? ??????? ?????????? ?????????? ???? ????????? — ??? ??????, ??? ?????? ?? ????????? ???????? ? ?? ??????????????? ??????????? ?????, ????????? ?????? ? ??????????? ?????????. ???????? ??????????? ??? ?????? — ?????????? ???? , ??????? ???????????? ??????? ?????????. ???, ? ???????? ???? — ??? ??????? ????? ??????? ?????, ????????????? ? ????????? ?????? ? ?????????? ????????? ??????, ??????? ???????, ???????????? ??? ???????, ?????????????? ???????. ???? ??????? ???????? ????????????? ???????? ??? ???????? ??????? ????? (?? 5 ? ????? ????). ???????? ???????? ??????? ????? ? ????? ???????? ????? , ????? ??????? , ??????? ????????? , ??????? ???????? ? ???? ?????? ??????????????? .

???? ?????? , ?????? , ????????? — ??? ??????????????? ??????? ??????? ? ????????? ????????????? ??????, ?????? ?????? ?????? ????????? ?????; ??????? ????????? ?????? ? ?????????????? ???????. ??? ???? ????? ? ??????? ???? ???? ??? ??????? ? ???????? ?????? ???????, ? ?? ?????? ?????? ??????? ??? ?????? ??? ????? ???????? ????? — ?????????????? ?????????? ?? ?????? ??? ??????? ????????. ???? ??????? ???? ? ??????? ???????????, ????? ?????????? ??????? ????, ??????? ?? ????????? ??????? ????????, ????? ? ???????? ??????? — ?????? ????????? ?????? ??????? ???????????? ???????????? ????????????? (????? ????? ??????? ?????? ?????????? 20—30 ?, ???????? ? ?????????????? ??????? 600 ?). ?????? ????? ????????? ?????? , ?? ??????? ????????? ????????????? ????????? .
?????? ????????????? ????? ?????????? ????, ????????????? ??? ??????????? ??????? ????? ?????????? ??? ??????? ???????? , ???????? ??? ????? «?????????? ????». ??? ???? ??? ??????? ??????????? ?????????? ???? ??????? ?? ????????? ?????? ????????; ????????, ??????? ?????? ? ????????????? ?? ?? ?????????? ??????? ??????? ????? ??????????? ??????? ????????? ?? ??????. ? ?? ?? ????? ??????? ???????? ? ????????? ????????? ??????????????????? ?????????????: ?????????? ?????????? ?????? ????? ???? ???????????? ? ????? ? ??????????? ??? ??????? ??????? ???????? ???? ???????? .
???????????? ?????????? ?????????? ????????? ????????????? ? ??????? ??????. ??????? ????? ?????????? ? ???????, ????????? ??????????? ????. ????? ?? ?????????? ???????? ????? ???????????? ?????????????? ? ??????? (?????, ???????? ??????, ????? ??? ? ?????). ??? ?????? ????? ?????????? ????? ?????? ?? ????????? ??????. ? ?????? ? ?????? ????????? ????? ????? ????????? ????? «??????? ????????» ? ??????????? ?????, ?????? ?????? (?? ?? ??????) ????? ??????? ????????????? ??????????? ??????? .
??? ????????????? ?????? ?????????? ??????? ???????? ????????? , ???????? ? ?????????. ???? ???????? ?????????? ???????? ?????? ? ?????? ????????? ???????????? ? ???????? ??????? ? ?????? ???????????? ???????? . ?????? ???? ? ????? ? ?????? ???????????? ???? ? ????????? ? ????? ????????? ??????, ??????? ?? ??????????? ? ?????? ????????? ? ?????????????, ??? ??????????? ?????? ??????????? ???????? — ????? ???? ????. ???????? ? ???????? ??????? ????????? ? ???????????? ??????? ?????? .
???????????? ??????????? ?? ????? ??????? ??????? ? ?????? ? ??????? ????????????? — ?????????? ??? ???????. ?????? ? ????? ???????? ???????? ???????. ???, ????? , ??????????? ?? ?????, ?????????? ?????, ????? «????????» (???????? ??????? ?? ????? ??????) ? «??????» (??????? ?????? ? ???????); ???????????? ?????? ????? ?????????? ???????????? ????? ????????? .

?????????? ????????????? ?????? ? ?????? ????? ????????????? ???????? ? ??????????? ????????? ??? ??????? ??????????? ?????? ? ???????????? ?????? ?? ????????????? ???????? ????? ? ?????? ?? ?????? .
?? ?????????????, ? ??????? ????????? ???? ????? ??????????????, ? ???????? ?????? ?????????? ???????, ?????? ????? ????????? ??? ???????? ????? ?????. ?????? ????????????? , ???????? , ????????????? , ?????? ??????? ? ?????? ; ????? ???????? — ??????????? ?????? ???????? ? ??????????? ??????? , ??????? ? ?????????? .
??? ??????? ????????? ????????, ???????? ???????? , ?????????? ??????????? ???????? ????, ???? ??? ??????. ??? ?????????? ??????????? ????? ????? ??????? ??????? ??????? ?????????, ????????? ? ????????????? ??????, ????????????? ?????? ????????? ? ??????: «????????» (?????), «????????????» ? «?????????????»; ??? ???? ???????? ?????? ???? ? ?????? ??????, ??????? ??? ????????? ????? ???????? ?????????. ? ????????? ??????? ?????????? ?????????? ????????????. ???, ? ????? ( ??????? ) ????? ????? ? ???????? ?????? ??????? ??????? ???????? ?? ????????? ???????, ? ????? ???????? ???? ? ????????? ? ????????? . ? ????????? ?????? (????????, ? ????????? ????? ) ? ?????? ??????????? ?????????? : ????? ??????? ?????? ? ?????????? ??? ???? .
???? — ???????????? ?????????? ???????? ????? ???????? . ????? ?????? ?????? ?????? ????? ????? , ??????? — ??????????? ?????????? . ?????? ?? ????????? ????? ???????????? ??? ?????????? ????? ( ??????????? ?????? , ??????? ????? ). ??? ?????????? ????? ???? ?????? ?? ??????? ????? ??????????? ????????? ????? ????????????, ? ?? ???????? ???????????????: ???? ????? ??????? ??????, ?????? ???????? ?? ??? ?? ?????? ??? ?????????? ?? ???? . ??????????? ?????? ??? ?????????? ????? ?? ??????? ???????? ????? ???????????? ???? ????????: ?? ????? ?????? ?????? ??????????, ???????? ??????????? ????????? ??????? ??????, ??????? ????? ?????? ??????? ????, ??????????? ????????? ??????; ????? ???????? ????? ? ????? ?? ?????????, ??????? ????????, ?????? ?????? ??????? ?? ????. ?????? ????????? ? ??????? , ?????????? ?? ??????? ?????? (????????, ????) .

???????? ?????? , ????????? ???????? ?????? ? ????????? ?????? ???? ???????? ??????? ?????????? ????????????, ??????? ????? ?????????? ?? ???????? ??????????????? ?????? ? ???????? ????? ?? ????? ??? ????????? ?????? ?????. ???????????? ???????? ??????? (?????? ?? 100 ? ????? ??????) ? ????? ?????????? ????????? ????? ?????? ????????? ???????? — ??????? (?????????, ???????, ???????). ????? ??????????? ????? ???????? ???????????????? ?????? ??????? ? ?????? ? ????; ??? ???? ? ?????????? ?????, ??? ???????, ??????????? ???? ?????-?????, ??? ???????? ??????? ??????? ?????? ?????????????? ??????????? ??????, ? ??????? ???????? ????????????? ????????? .
??????? ????? ?????????? ??????? (5—30 ??????); ??? ?????? ???????? ?????-??????, ?????? ??? ?????????? ??????? ??????, ?????????? ???????? ????? ? ?????????. ????????????? ????????? ????? ??????????? ? ??????? ?????????? ?? ??????, ?????? ????? ???? ?? ????? ???????. ? ????????? ?? ????? ????? ???? ??? ???????????, ?? ?? ??????, ?, ? ??????? ?? ??????????, ?? ???????? ???????? ??????: ????????? ????? ? ?????? ????? ???????????? ????????. ???????? ????? ???? ?????? ???????, ???? ?????????? ???????; ?????? ???????? ????? ??????????, ????????? ? ?????, ?? ?????????? .

? ???????? ??????? ????? ????? ????????? ????? ????, ???????? ? ??????? ?? ????????. ???????? ??????, ????? ???????? ????? ? «????????? ?????» ????????? ???????? . ????? ????????? ?? ??????? ?? ???????? ( ?????? ??? ???????? ), ? ???????? ????????? ???? ? ???????? ?????? ? ????????? ????????? ???????; ????????? ??????? ??????????? ?? ???????? ?????? ? ????????. ????? ????????? ???????? , ?????? ???? ????? , ???????? ? ??????????? ? ? ????????? ????? ?????????? ?????, ???? ?? ????????? ??????? ??? ?? ?? ???????? ??? ????. ??????????? ??????? , ???????? ??????? ??????, ???????? ???????? ? ??????? ???? ????? ????? .
????????????? ????????? ????? ????????? ? ??????? ??????? ????? ???????? ????? ????? ?????????????, ?????????? ??????? : ?????? , ?????????? , ????????? ??????????????? ( ??????? , ?????? , ??????? , ????? , ???????? ? ??.); ????? ?????? ????????? ????? ???????? . ????? ????????, ???????? ? ?????? ???????????? ? ?????????? ???????, ????? ???? ?????????????? ? ???????? ???????, ?? ??? ???? ??????????? ?? ????????? ??????????? ????????? ?? ?????? ????? ??????????? . ??????? ????? ???????? ????????? ???????? ??? ????? ????????; ???????? ??????? ???????, ????????????? ?????? ?????? ??????, ?????????? ??? ?????????? . ???????????? ? ???? ????????? ???????? ??????????? ???????????? : ? ????????? ??????? ?? ??? ??? ?? ???????? ??????? ???????????? ?? 20 ??? ?????? — ????? ??????? ?? ????? ????????? ????????????? .

????? ???? ????? ??????????? ????????????? ?????????? ????????? ??????? ???????? ?????????? , ??? ??????? ?????? ????? ? ????????????? ????????? ???? ????????? ????????? ?????, ???? ???? ??????? ??????????? ??????????? ? ????????????? ?????? ??????????????? ????? . ????? ?? ??????????? ??????????? ????????, ?????? ?????????? ? ?????????????, ???????? ??????????? ??????????? (????????? ??????????? ? ??????? ????? ????? ? ????? ????? ????). ?? ????? ?????????? ?????????? ????? ???????? ??????? , ? ?????? — ???????? , ? ??????? ???????? ??????? ?????????? ???????? ? ??? ???? ?????? .
???????, ??????????? ?????????????? ??? ?????????? ?????????? ????????? ???????? ?????? ?????????? ?? ???????????????????, ? ??????????????? ??????? . ? ?????? ?????????? ??????? ???????? ???????? ?????????????? ???????, ????????? ???????? ?????????? ???????? (? ???? ????? ?????????? ??????????? ???????? ???????? , ??????????? ? ?????? ??????? ), ??????????? ????? ??????????? ?? ??????????????? ?????? (????? ??????????, ??? ? ??????????? ????????????? ?????????? ??????? ?? ????? ?????????????? ??????? ?? ???????? ?????? ?????? ???????????, ?? ??????????? ????? ??????? ??????? ????????????? ????????? ???????? , ???????? ???????????????? ) .
? ???? ??????? ?????????????? ??????? ????????? ????????????? ? ????????????? ????? ???? ????????? ? ??????????????? ?????? ????????? ????? ?????????. ???, ??? ??????????? ?? ?????????? ?? ??????? ????????? ???????? ??????????, ??? ????? ?? ?????? ??? ??????????????? ??????? ??????? ????????? ?? ????? ? ???? ????? ?????? ? ????? ?????? , ????? ???? ????? ?????????? ?????????? ???????? ? ?????? ??????, ?????? ?? ??????? . ? ???????????????? ??????? ??????? ???????? ??????? ???????????? ??????? ? ????????? ????????????? ???????? : ????????????? ?????????, ??? ?????? ???????? ??????? ??????? ?????? ???????? ? ??????? ????????????? ? ???? ?????????? ?????????. ?????????, ??? ???????????? ????? ??????? , ??????? ????? ????? ?????????? ?????????, ??????????? ????????? ?? ?????????? ????????, ?? ??????? ?????? ????? ???? ?? ? ????????? .
??????, ????????? ???? ????????????? , ??????? ??????? ? ???, ??? ? ???????????? ????? , ????? ????????? ? ????? ?????????? . ? ?? ?? ????? ? ?????? ???????????? ???? ???? ?????? ?????: ???, ???? ???????????? ????? ???????? ????? 8 ???????? ????????, ?????????? ??????? ???????? ? ???? «???????» («????????!», «???????!», «??????», «?????», «????????????») . ? ????? ???????? ??? ????? 30 ???????? ????????, ??????????? ??????? ? ??????? (????????, ??????????? ?????? — ???? ?????????????) .

??????? ????????? ????????????????? ????????????? ???????? ???????????????? ????????. ? ???? ?????????? ??? ?????????? « ???????? ???????? » ??????? ??????? ???????? , ?????? ? ?????? ?????? ????????????? ?????; ? ???????? ?????????? ????????? ????? ????? ?????????? , ?????????? ??????????? ???????? ??? ??????????? ?? ?????? ??????????. ? ?????????? ???????? ???????? ???? ???????? ????????? ???????? ??????, ?????? ??? ????? ?????????????? ?????????? ????? ????? ? ?? ????????? (??????????? «???? — ?????» ???? ??????? ??????? ???????????? ?? ?? ???????? ??????), ? ????? ??????????? ????? ? ?????????? ???????? . ??????? ???? ? ?????? ???????? ????? ???????? ????? 500 ??????? .
??????????, ??? ???????????????? ???????? ???????? ???????? ??????? ????? ????????? ? ???????? . ???, ?????? ????? , ?????????? ? ?????? «????????? ???????» ? ???????? , ??????? ?????? ????????? ????? «?????? — ???????? — ????» ?????????????????: «???? — ???????? — ??????». ???? ??????? ?????????? ??? ?????????, ?? ????? ??? ???????? ?????, ?????????? ???????????????? . ??????????? «???????? ????????» ???? ????? ? ???, ??? ????????? ???????????????? ??????? ?????????? ??? ????????? ???????????? ?????? ?? ?????? ??????? ?????? ??? ????? .
????????????? ??????? ?????? ??????? ??? ?????????? ????? . ? ?????????? ???? ? ????? ??? ???????? ????????????? ???? ??? ????????? ???????????? ???????????? ??????, ??? ? ?????????? ????????. ????? ??????????? ????????????? ?? ??????????? ? ???????? ? ???????? ???????? ????????? ????? , ? ?? ???? ?????????? ?????? ?????. ?????? ????????????? (???????? ??????? ? ???? ) ????? ? ???? ???????? ??????????? ????????? ????????? ? 100—200 ???????? ?? 1 ?? ; ??????????? ?? ??????? ???????? ? ?????? ?????? ?? ????????? 1—5 ?? 1000 ?? .
??????????? ??????? ?????? ??????? ????? , ????????????? ????????? ??????? ?? ???????? ???????? . ? ????? ???????? ????????? ??????? , ? ? ??????? ? ?????? ?????????? ???????? ????????? , ??? ???????????? ????????. ?????? ???? ???????? ?????????????? ? ??????????????? ????? ? ?????? ??? ?????????? ?????????????: ???? ???? ??? ? ????????? ? ????? ? ?????? ?????? ??????? ? ??????? ( ???????? , ?????? ) ??? ???????????? ? ?????? ????????, ??? ? ??????????? ???????? ????????? ??????, ???????? ? ?????? ??????. ?????? ?????? ?? ?????? ??? ??????????? ????? ??????????????? ????? ????????? ????? ????????? , ? ? ????? ?????? ? ?????? ????????? ???? ??????????: ??????????????? ??? ?????????? ???????????? ???????? ?????? .
???????????? ????????????? ?? ?????? ?????????? ???????????? ????????????? . ???, ? ???????? ? ?????? ????????? ??? ??????????? ?????? ???????? ( ??????? , ???????? ) ?????? ??????????? ???????? ????? ????????- ????????????? ? ?????????? ????? ????? ????????????-???????? , ??? ???????? ???????? .
????????? ????????????? (? ???????? ??????? ) ????? ???????????? ?????????????????? ????????, ?????? ????????????? ???????????? ????????, ??????? ??? ???????? ? ?????? ????????????? ( ???? , ????????? ? ??.) .

?????? ??? ????????????? ??? ??? ???? ?????, ????? ? ???????? ????????????? ????? ?????????? ??? ???????????? (????????? ?????????? , ?????????? ????????? ??????? ), ??? ? ????????????? (??????????? ???????? , ??????????? ?????????? ????? , ?????????? ???????? ) ???????. ??? ??????????? ??? ?????????????? ????? ????? — ?????? ? ?????????? ??????? ? ??????? , ?????????? ???????? ? ?????????? ???????? ??????? ( ????????? , ?????????? , ) ? ??????? , ?????????? ? ????????? — ??????? (?? ??? ??????? ????????) ?? ????????? ??????????? ??????? (10—12 ???. ??? ?????) .
? ???????????? ????? ??????? ????? 100 ????? ??????? ?????????????, ? ??? ????? 33 ???? — ?? 1700 ?., 36 ????? — ? XVIII—XIX ??. ? ????? 40 — ? 1-? ???????? XX ?. ????? ???: ??? (????????? ????????????? ????? ?? ????????? ? 1627 ?.), ?????????? ?????? (????????? ?? ??? ????? ? 1768 ?. ?? ??????? ??????? ), ?????? (????????? ?????? ?????? ? 1883 ?. ? ????????????? ???????? ), ?????? (????????? ????? ??????? ?????? ? ?????????? ???????? ? ????? 1880-? ??.) .
? ?????? XXI ???? ??? ??????? ??????????? ??-?? ?????????????? ????? ????????? ???? , ????? ? ??????????? ?????? ? ??????? ?????? ?????? , ????? ? ??????? ? ??????? ???? ; ???????? ?????? ? ???????? ??????? ??????? ? ????? ??????? ??????????? ??????? ??????? ? ???????????? ??????? ; ??????????? ????? ?????? ????????????? ??????? ????????? ?? ????? ???????????? ??????????? ????? .


??? ?????????? ?????????????? ???????????? ????????????? ????? ???????????????? ??????????????? ? ???????????? ??????. ?????? ? ???? ????? ?? ?????? ????? ???????? ????? ???????? ? 1868 ???? ??????? ?????? ?? ?????? ? ???????? ??????-??????? ??????? ?? ?????? ???? ? ?????? ? ?????? .
????? ?? ????????? ??????????????? ??? ????? ??????????? ???????????? , ? ??????? ??????? ?????? ?? ?????????? ? ?????????????? ??????????? ?????? ????????. ?? ?????????? ?????????? ??????? ?????? ??????? ?????????? — ?????????? «???????-????» — ??????? ?? ?????????? ?????? ?????? ?. ?. ?????-???? ? 1889 ???? .
? 1966 ???? ???? ?????? « ????????????? ??????? ????? », ? ? 1978 ???? — « ??????? ????? ???? »; ? ??? ???? ????????? ?????? ?????????? ? ?????? ????? ? ???????? ? ?????????????? ?? ?? ?????? (? 1983 ???? ??????????? «??????? ????? ?????» , ? ? 2001 ???? — « ??????? ????? ?????? » ). ?? ????????? ?? ??????? 2014 ???? ? ????????????? ??????? ????? ??????? ?????? ?? ??????????? 5506 ????? ?????????????, ?? ??? 77 ????????? ????????? ????????? (????????? EX), 2 ???? ???????? ????????? ? ????? ??????? (????????? EW), ? ??? 1143 ????????? ??? ??????? ????????? ??????? ????? (????????? LR, VU, EN, CR) . ? ??????? ????? ?????? ???????? 74 ???? ????????????? .
????? ???????????? ????? ???? ?? ?????????? ???? ????????????? ???????????? ????????. ??? ?????????? ?????? ?????? ????????, ????????? ?? ???????? ????? ????? ??????? ????????? , ? ??? ????? ????????? ????????? ????? ????: ? ??????????? ???? — ???????? , ????? ???? , ?????? , ??????????? , ????? ; ? ????? ???????? ??????? — ????? , ?????? , ????? , ?????? , ????? ?????? , ??????? , ?????????? ???????? , ???????? ??????? , ???????? ???? . ??? ????? ??????? ?????????? ????, ????, ???????? ????? ? ???? ??? ???????????? ?????? .
?? ?????? ????????????? ????????? ????? ?? ????? ????????????? ????????? ? ??????????? ???????? ????? ???? ??????? ?????????, ?? ????????? ???????????? ?????????? ?? ??? ??? . ? ????????? ????? ?????????? ?????????? ?????, ???????? ????????? ??????????? ? ?????????? ?????, ????????? ? ??????? ?????? , ???????? ? ????????????????? .
??????????? ?????????? ????? ??????? ??????????? ????????. ????? ?? ???????? ????? : ?????? , ??????? ? ????????? ??????? , ?????? , ????? — ??????? ???????? ??????? ????????????? ????????? ????????? ?????? , ??????? , ????????? , ??????? . ???? ?????? ???????? ????? ? ?????? ??????? ?? ?????????? ????????????? ?????, ?????? ??????? ?????? ?????????? ?????????????? ???????????? .
? ????? XIX ???? ???????? ? ???????? ?? ???????? ???????????? ???????? ????? ??????? ????????? — ?????? ???????????? . ??? ???????????? ?????????? ?????? ?????? — ?????? , ????? , ????? , ?????? , ?????? , ???????? — ?? ??????????? , ??? ???????? ??????????? ? ???????? ??????? . ?????? ????? ?????? ?????????? ???? ???????????? ? 1895 ???? ? ?????? ? ??????????-??????? ???????? , ????? ??????? ? ??????? .
?? ?????? ?????? ?? ?????? ???????? ????????? ?????? ? ??????? , ? ????? ????? .
???????? ????? ???????????? ????? ???????? ????????? ? ?????????????, ? ?? ??? — ? ????????????????? , ? ?????? ? ?????? ????????????? . ????? ?????? ??????? ?????????? ????????????? ????????????? ( ???????? ?????? ), ???????? ( ??????? ?????? ), ?????? ( ?????? , ????? , ?????? ), ? ??????????????? ( ?????? , ???? ) . ????????????? ? ?????????????? ??? ???????????????? ???????????? ?????? ???? ? ????????? ???????????? ??????? ???????? ???????? ?? ????? ????? ???????: ???, ? ???????? ????? ? ?????? ??????????? ??????????? ??? ? ??????????????, ??????????? ?????? ??????????? ?? ???????? ? ???????? ???? ????????? ?????? ????? ? ????? . ???????, ?????????? ?????? ??????? ????? ?????? ??? ??????????? ? ?????? ???????????? ?????? ???????? ?? ??????; ??????? ??????????? ????? ???????? ? ??????? ? ????? ????????????? ?? ??????? ??????? ?????????????? .

?????? ???????? ???????? ????? ?????? , ???????????? ??? ? ??????? ????????? — ?????? ? ?????? ???, ? ??? ??????????????? ?????????????? ? 2008—2013 ??. ?????????? ??????? ??? ?????????? ???????? ?? ?????? ???? ? ??????? , ??????? ??????? ???????? 31,7 ???. ???, ? ?? ?????? ??????????? ?? ????? , ? ??????? ??????? ??????????? ? 33 ???. ??? ). ????????????? ??? ??????? ?????????? ????????, ?? ????? ????? ?????????????? ? ?? ?????. ? ?????????? ??????? ????? ????? ????? , ?????????? ????????? ? ?????? ?????????? ??????, ????????? 350 ; ??? ???? ????????????? ?????????????? ????????? ???????? 343 ?? ??? . ??????????? ??????? ???? ?????????? ????? . ?????? ???????, ??? ??? ??????? ???????? ????????????? ???????? ?????? ( ??????? ?????? , V ??????????? ?? ?. ?.), ?????????????? ????? ??? ?????? ????????? ??? ??????? ?????? ? ??????? ?? ???????? . ?????? ??????????? ? ????? ??????? ??????? ??????? ( ???? , ????????? ) ???????? ??????? ; ??????? ??? ??????? ???????? ???? ??? ????????????? ?????? ??????? ????????????? .

????????????? ???????? ???????? ? ???????? ???? ?? ?????? ???????? ? ??????? ? ??????? ? ????????????? ???????????? — ?????? ? ??????????? , ????? ?? ???? ?????????? ???? ????????????? ????????? . ???????????? ???????? ????? ???????? ?????????? ????, ??????, ?????? ? ????, ?????????????? ??? ???????????? ? ???????? ????????. ??????????????? ???? ??????????: ???? (??????? ????-???? ??????? ? ???????? ????? , ????? X—IX ??????????? ?? ?. ?.), ???? (??????? ?????-???? ? ???????? ????? , IX ??????????? ?? ?. ?.), ?????? ( ? ???????? ????? , VIII ??????????? ?? ?. ?. ), ?????? (????????? ?????? ?? ???-??????? , ????? VIII ??????????? ?? ?. ?.; ????? ?????? ??????? ?? ??????-???? ? ?????? ???? ???? ?????????? ?? ???????? ????? ?????? , ?? ???????? ???????? ?????? ?? ?????????????, ? ?? ??????? ?????????? ). ? ????? ???????? ??????? ????????? ????????????? ?????? (????? ? ????????? ????????? ?????? , VI ??????????? ?? ?. ?.), ???? ( ????? , 1-? ???????? V ??????????? ?? ?. ?.), ?????????? ( ??????? ???? , 2-? ???????? V ??????????? ?? ?. ?.) ? ??????????? ???????? ( ?????? , IV ??????????? ?? ?. ?.) .

????? ???????? ????? ????? ?????????????. ? ????? ? ????? III — ?????? II ??????????? ?? ?. ?. ?????????? ??????? (? ???????? ???????? ???? , ???????????? ??? ??????? ????????) ? ??????? (?????? ??????? , ?????? ??????); ? I ??????????? ?? ?. ?. ????? ?? ???? ?????????? ??????? ?????? , ??????? ? ?????? ??????? ???????? ?????????? ???? . ? ????? ? ???-????????? ???? ????????? ???? ????????????? ???????? ???????? ?????: ????????? ?????? (???????????????? — ? III ??????????? ?? ?. ?.), ???? (????? ?????? — ???? ) ? ??????? . ?? ?????????? ?????????? ??????? ? III ??????????? ?? ?. ?. ??? ????????? ?? . ? ???????? II ??????????? ?? ?. ?. ? ?????? ????????? ???????????? ????? ??????? ?????? — ?????? , ? ? I ??????????? ?. ?. ?? ??????? ??? ????????? ?????? . ? ?????? ??? ????????? ???????? ????? , ?? ????? ?? ?????? ????: ???? ?????? ???????? ?????? ???????????? ??????? ???????? (III ??????????? ?? ?. ?.), ?????? — ??????? ????????? ???? (I ??????????? ?? ?. ?.) .
???????, ??? ????????? ????????????? ??????? ??????? ?? ????, ??????? ???????? ???????? ? ???????? ???????????? ? ??????. ???, ????????? ???? , ??????? ???????????? ? ??????? ????? ? ???-????????? ???? ??? ?????????? ????????????? ?????, ?? ????????? ? ???????? ????????, ????????? ? ?????? ?? ??????????? ?? ???????????? .
????? ??? ????? ????????????? — ???? , ????? , ??????? , ??????? ?????? , ??????? , ?????? , ????????? ???????? — ???????????? ? ???????? ???????????? ???????? ??? ?????????? ???????????????, ???????????? ? ??????????? ???????????? .
? ????????? ???? ??????????? ??????? ???????????? ?????????????, ???????? ???? ???????? ???????? . ?????? ??????? ???????? ???????? ( ?????? , ????? , ??????? , ??????? ?????? ), ???? ?????? ???? ???????????? ????? ? ???? , ?????????? ? ?????????? ???????? , ??????? , ???? ? ??.
??????? ???????? ???????? ????????????? ???????? ??? ? «???? ????????» ?????????? , ??????? ? ?????????? « ??????? ???????? », « », « ? ?????? ???????? » ?????? ???????? ? ?????? ????? ????????????? ??????????????? , ?? ???????? ? ?????? ?????, ???????????? ??????????? ? ???????? ?????????????. ?? ?? ????????? ?? ? ?????? ??????, ? ???????????? ? ??????? ???? ?? ???? ?????????? ?? ???????? ????? «???????? ? ??????» (?? ???? ???????????): «??????????? ????????????» ? «??????????? ????????» ( ???? ) .
? ??????? ???? ??????? ? ???????? ????????? ???? ????????, ?? ????? ?????? ? ??????? ?????? ??????? . ????? ?????????????? ???????? ???????? ????????????? ?????? ??. ??? ? ?. ?????? , ? ??????? ???? ???????? ?????? ??????????? ?????????????? ? ????????????? ???? , ??????? ????????? ???? ? ????? ?????? Mammalia . ??????? ???????? ????? ?????? ?. ????? ? ?. ??????? ????-????? , ??????????????? ??????? ????????? ? ???????????????? ??????? « » ?????????, ? ? ??????????? ????????????? ???????????????? ????????? ??????? ??????. ????? ??????? ?????? ????????????? ?????????????. ????? ? ??????????? ????????????? ?????? ????? ?. ??????? , ?. ??????? , ?. ???????? , ?. ???? . ?????? ??????????? ????????????? ???????? ?. ?. ????? ? ?. ?. ??? .

????? ? ???? ????? ?. ??????? « ????????????? ????? » (1859) ??????? ?????? ??????????? ???????? ????????????? (??????????) ?? ?????? ????????????? ????????????? . ???????? ???? ? ???? ??????????? ??????? ?. ?????? , ?. ?????? , ?. ??????? , ?. ?. ??????????? , ? ????? ????????????? ???????????? ??????????????? ?????, ???????????????? ?? ???? ????????????? ????? ???????????? ??????? : ?. ??? , ?. ?????? , ?. ??????? , ??????? ???????? ?????? ????? ?????????? ??. ?. ??????? ? ?. ???????? . ? ?????????????? ? 1910 ???? ???????? ????????????? ??????? ? ??????? ??? ?????? ??????????????? ????????? ???????? ???????????? ??????????? . ??????????? ?? ???????? ??????? ???????????? ?????????????, ?. ?. ??????? , ?. ?. ?????? , ?. ?. ?????????? ???????? ?????? ??????????? ??????????????? .
? XX ???? ?????????? ?????????? ??????? ??????????????? ????????????. ??????? ????????? ??????? ???????? ?????????????, ?? ???????? , . ?????????? ?????????? ??????? ??? ??????? ?????????? ? ?????????? ???????? ( ???????????? , ???????????? , ?????????????? ? ??.).
???????? ?????? ? ??????? ??????????? ????????????? ????? ????????? ??????? ???????? (1945), ??????????? ?? ????????? ???????????? ?????????? , ? ??????? ???????? (1975; ????? ???????????? ????????????????) — ?????? ???????????? ??????? ?????? ??????????????? ????? . ? 1960-? ??. ? ????????????? ??????? ??????????? ????????????? ?????? ?????? ???????????? ???????????? . ??????? ???????????????? ??????? ????????? ?? ?????? ??????? ??????? ?????? , ? ????? ????????? ? 1977 ???? ??????????? ??????? ?????????????? ??? — ? ???????? ?? ??????? ???????????? ??????????????????? . ? ?????????? ?? ?????? XX—XXI ????? ????????????? ? ??????????? ?????? ??????? ????????????? ???????????? ???????????? ????????? .

? 1996 ???? ?? ???? ????????? ???? ????? — ?????? ????????????? ?????????????. ????? ???? ???????? ??????? ???????????????? ???????????? : ?? ???????????? ?????? ?????? ???????? ???? ???????? ????, ????????? ??? ? ????? ?????? ???? ? ???????????? ????????? ?????? ????????????? ????????????, ????? ???? ?????? ?????? ????????. ?? ?????? ???????? ?? ?????????????? ? ????? ??????? ????, ??????? ? ?????? ????? . ???? ????? ??????????? ??? ???????????? ? ?????? ????????????? . ? ?????????? ??????? ???? ??????? ??????????? ????? ?????????????, ??? ???? ? ?????? (1998), ???? (1999), ?????-????? , ???? ? ?????? (2000), ????? ? ?????? (2002), ?????? , ??? ? ????? (2003), ?????? ? ??????? (2005), ????? (2006), ?????? , ???? ? ????? (2007), ??????? (2010) ? ??.
? ?????? XXI ???? ???????? ???????? ??????? ? ?????????????? ??????? ????????? ????? ?????????????, ????? ??????? ?? ????? 2013 ???? ????????? ????? ??????. ????? ??? — ??????? (2001—2006 ??.) , ??????? ???? (2002) , ?????? (2005) , ???????? (2005) , ????? (2007) , ??????? ??????? (2007) , ??????? (2007) ? ??????.
| ?????? ????????? |
|
??????
????????
(Chordata)
|
|||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ??????????? (Acrania) | |||||||||||||||||||||||||
| Olfactores |
|
||||||||||||||||||||||||
|
† —
????????
??????, * —
????????????????
??????
|
|||||||||||||||||||||||||







