Вымышленные черепахи
- 1 year ago
- 0
- 0

Черепа́хи ( лат. Testudines ) — один из четырёх современных отрядов пресмыкающихся . Характерной особенностью строения черепах является костно-роговой или костно-кожистый панцирь, состоящий из двух частей: карапакса и пластрона , и служащий основной защитой от врагов . Многие виды способны втягивать под панцирь голову, хвост и конечности. Длина от 12 см до 2 м. Зрение и обоняние хорошо развиты, слух слабее. Челюсти лишены зубов и покрыты роговыми пластинками в виде клюва. Шейный и хвостовой отделы позвоночника подвижные, остальные неподвижно срастаются с карапаксом. Возникновение панциря эволюционно вызвало перемещение поясов конечностей под рёбра (исключительный случай среди позвоночных).
Наземные виды черепах распространены на территории всех материков, кроме Антарктиды и на многих островах. Морские виды широко распространены в тропических и субтропических зонах, реже — в бореальных районах мирового океана. Могут обитать в озёрах, реках, болотах, жарких пустынях, тропических лесах, на горных склонах, на культурных землях, морских побережьях, в океанах. Большинство видов ведёт полуводный образ жизни. В холодное и засушливое время года могут впадать в спячку.
Сухопутные и морские — преимущественно растительноядные; пресноводные виды — обычно плотоядные: могут питаться рыбой, земноводными, беспозвоночными. Спариваются обычно на суше, где и откладывают яйца (от одного до нескольких сотен штук). У некоторых видов может быть до 3 и более кладок за год. Яйца шаровидные или эллиптические покрыты у большинства сухопутных видов известковой скорлупой, а у морских и некоторых пресноводных видов — кожистой оболочкой. Инкубационный период обычно 2—3 месяца (у слоновой черепахи 6—7 месяцев). Половозрелость наступает в возрасте 2—3 лет. Рост неограниченный, но у половозрелых особей замедляется. Продолжительность жизни до нескольких десятков лет, иногда до 150 лет.
Отряд включает около 328 современных видов, группируемых в 14 семейств и два подотряда . Ископаемые остатки черепах прослеживаются на протяжении более 220 миллионов лет.
Несмотря на то, что традиционно черепахи рассматривались как анапсиды , все генетические исследования подтвердили гипотезу о том, что черепахи — это диапсиды с редуцированными височными окнами; некоторые авторы помещали черепах в группу лепидозавроморф , хотя все более поздние исследования подтвердили их положение внутри группы архозавроморф .
Черепахи распространены в тропической и умеренной климатических зонах почти по всей Земле. Экологически их подразделяют на морских и наземных , а наземных, в свою очередь, на сухопутных и пресноводных .
Многие виды черепах служат пищей человеку , но мясо некоторых может быть ядовитым . Также многие виды черепах находятся под угрозой вымирания разной степени и охраняются . Черепаха является распространённым символом в культуре многих народов .
Изучением черепах занимается герпетология .


Вопрос об эволюционном происхождении черепах остаётся открытым. Морфологические отличия в плане строения между черепахами и другими наземными позвоночными животными привели к тому, что в качестве предков либо близких родственников черепах рассматривались многие группы древних амниот . В современной научной литературе существует несколько филогенетических гипотез, описывающих вероятное эволюционное положение черепах в систематике высших позвоночных. На основании остеологических характеристик, черепах чаще всего рассматривают в качестве примитивных амнион — парарептилий , сближая их с диадектами , проколофонами или парейазаврами . Иногда их предками рассматривают — ранних амниот с неясной систематикой. Данные молекулярной систематики указывают на то, что черепахи могут являться одной из групп диапсидных рептилий . Диапсидное происхождение черепах также подтверждают и некоторые их морфологические черты, что позволяет ряду исследователей сближать их с завроптеригиями .
Условно их предками считались пермские котилозавры , а именно ( Eunotosaurus ) — небольшие, похожие на ящериц животные с короткими и очень широкими рёбрами, образующими подобие спинного щита . Но также встречалось мнение, что черепахи произошли от особой группы парарептилий — потомков дискозаврисков (амфибий) . Последние филогенетические исследования указывают на то, что черепахи являются диапсидами с редуцированными височными окнами и являются сестринской группой по отношению к архозаврам, в наше время представленным крокодилами и птицами . Таким образом, черепахи обособились значительно позже, чем считалось ранее.

Ископаемые остатки древнейших черепах длительный период времени были известны исключительно из отложений позднего триасового периода (210 млн лет назад) Германии, откуда был описан Proganochelys quenstedti , изученный по нескольким хорошо сохранившимся скелетам. Она характеризовалась полностью сформированным панцирем и наличием примитивных черт в строении черепа (полная крыша, нёбные зубы) и скелета (туловищный панцирь с большим количеством щитков и пластинок, крупные остеодермы на шее, конечностях и хвосте) и на основании этого была выделена в особую базальную группу черепах — Proganochelydia. С конца XX века география находок триасовых черепах значительно расширилась: их окаменелости были обнаружены в Таиланде , Аргентине , США , Гренландии . Наиболее знаковым открытием является описание наиболее древней ископаемой черепахи Odontochelys semitestacea из верхнетриасовых морских отложений Юго-Западного Китая. Она характеризуется комплексом уникальных и крайне примитивных признаков: челюсти с зубами, пластрон хорошо сформирован, карапакс недоразвит, рёбра (костальные пластинки) слабо расширены и не контактируют между собой, маргинальные пластинки отсутствуют, пластинки над позвонками отделены от остистых отростков позвонков. Среди необычных чертам вида следует отметить относительно большое количество фаланг пальцев и прикрепление рёбер в средней части тел позвонков, а не между ними, как у других черепах. Данный вид показывает промежуточную ступень эволюции, переходную между типичными тетраподами и типичными черепахами . Даже самые ранние позднетриасовые формы черепах характеризуются наличием и типичным строением панциря.
В позднем меловом и кайнозойском периодах представители семейств и Podocnemididae получили широкое распространение в Северном полушарии из-за своего прибрежного образа жизни . Самые древние известные черепахи с мягким панцирем и морские черепахи появились в раннем меловом периоде . В том числе в меловом периоде возник крупнейший известных вид черепах, известный по окаменелостям из Южной Дакоты (США) — морской вид архелон ( Archelon ischyros ) — крупнейший экземпляр весил 2,2 т, при длине тела 4,6 м от головы до хвоста .
В Азии черепахи эволюционировали в эоцене . Поздняя выжившая группа из клады тестудинат — меиоланииды существовала в Австралазии в плейстоцене и голоцене . Среди них были крупнейшие сухопутные черепахи рода миолания ( Meiolania ) с длиной панциря до 2 м. Они имели огромный, практически одной длины с панцирем, мощный хвост, усаженный двумя рядами костяных уплощённых шипов, а на концах их треугольных черепов располагались длинные притупленные «рога», направленные назад и вбок .

Точное происхождение черепах оспаривается. Считалось, что они были единственной сохранившейся ветвью древней эволюционной категории Anapsida , которая включает такие группы, как проколофониды и парейазавры. У всех анапсидов в черепах отсутствует височное отверстие, в то время как у всех других живых амниот височные отверстия имеются . Позже было высказано предположение, что анапсидоподобные черепа черепах могут быть результатом обратной эволюции, а не анапсидного происхождения . Ископаемые остатки свидетельствуют, что ранние базальные черепахи обладали небольшими височными отверстиями .
Некоторые ранние морфологические филогенетические исследования помещают черепах ближе к Lepidosauria (ящерицы и змеи), чем к Archosauria (крокодилы и птицы) . Напротив, несколько молекулярных исследований помещают черепах либо в состав архозавров , или, чаще всего в качестве сестринской группы по отношению к существующим архозаврам , хотя анализ, проведенный Тайлером Лайсоном и его коллегами (2012) вместо этого восстановил черепах как сестринскую группу лепидозавров . Дата эволюционного разделения предков черепах, птиц и крокодилов оценивается в 255 миллионов лет назад во время пермского периода . Николас Кроуфорд и его коллеги (2012) также обнаружили, что черепахи ближе к птицам и крокодилам, проводя филогенетическое исследование ультраконсервативных элементов (UCE) в геномном масштабе .
Используя предварительные (незавершенные) последовательности геномов зелёной морской черепахи и китайской черепахи с мягким панцирем, Чжуо Ван и его коллеги (2013) пришли к выводу, что черепахи являются сестринской группой крокодилов и птиц . Внешняя филогения черепах показана на кладограмме ниже .
| Sauria |
|
||||||||||||||||||||||||
Современные черепахи и их вымершие родственники с полным панцирем относятся к кладе Testudinata . Время существования последнего общего предка современных черепах ( бокошейных и скрытошейных ) оценивают примерно в 210 миллионов лет назад (поздний триас) . Роберт Томпсон с коллегами (2021) отмечают, что современные черепахи отличаются низким для настолько древней группы разнообразием. Согласно их анализу, видовое разнообразие было стабильным, за исключением одного скачка на границе эоцена и олигоцена около 30 миллионов лет назад и крупного регионального вымирания примерно в то же время .
Кладограмма показывает внутреннюю филогению Testudines вплоть до уровня семейств. Анализ, проведенный Томпсоном и его коллегами в 2021 году, подтверждает ту же структуру вплоть до уровня семейств .
| Testudines |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Крупнейшей среди ныне живущих черепах является кожистая черепаха ( лат. Dermochelys coriacea ), чья длина панциря достигает 2,5 м, а масса тела может превышать 900 кг. У крупнейшей достоверно измеренной кожистой черепахи полная длина тела составляла 2,6 м, размах передних ласт — 2,5 м, а масса — 916 кг . Пресноводные черепахи, как правило, мельче морских, однако среди представителей наиболее крупного вида Pelochelys cantorii известны экземпляры с длиной тела до 2 м. Это значительно превышает размеры тела самой крупной североамериканской черепахи ( грифовая черепаха ( Macrochelys temminckii )), чья длина тела достигает 80 см, а масса — 113,4 кг . Гигантские черепахи родов сухопутные черепахи ( Geochelone ), миолания ( Meiolania ) и других были широко распространены по всему миру в минувшие периоды; их ископаемые остатки известны в Северной и Южной Америке, Австралии и Африке. Они вымерли одновременно с появлением человека, и предполагается, что они были истреблены охотившимися на них людьми. Единственные сохранившиеся до наших дней гигантские наземные черепахи обитают на Сейшельских и Галапагосских островах; максимальная длина тела слоновой черепахи равна 187 см , а масса — около 400 кг . Самой большой из известных науке черепах считается жившая в меловом периоде морская черепаха архелон ( Archelon ischyros ) . Размер одного из обнаруженных скелетов этого вида достигает 4,5 метров, а предполагаемая масса этой черепахи — до 2,2 тонны . Однако, крупнее архелона может оказаться вымершая позднемиоценовая—раннеплиоценовая пресноводная черепаха Stupendemys geographicus, с крупнейшим известным карапаксом длиной 3,3 метра и указывающим на общую длину около 5,25 м, а также вес около 6 тонн .
Самой мелкой черепахой является капская крапчатая черепаха ( лат. Homopus signatus ) из Южной Африки . У самок длина карапакса достигает 11 см, а масса — 241 г. Самцы существенно мельче, и длина карапакса у них не превышает 9,7 см, а масса 124 г . К мелким черепахам также можно отнести представителей двух родов американских черепах — замыкающиеся черепахи ( Kinosternon ) и мускусные черепахи ( Sternotherus ), чей ареал простирается от Канады до Южной Америки. Длина панциря многих видов этих родов меньше 13 см.
Большинство черепах всю жизнь обитает на поверхности земли, и их глаза расположены по бокам головы и направлены книзу. У некоторых водных черепах глаза расположены близко к верхней части головы. Эти черепахи могут прятаться от хищников на мелководье, полностью погружаясь в воду и оставляя на поверхности лишь глаза и ноздри. Кроме того, около глаз у них располагаются слёзные железы, выделяющие солёные слёзы. Таким образом черепаха выводит излишек солей, который попадает в её организм вместе с выпиваемой водой.
У черепах имеется твёрдый клюв, который они используют для откусывания пищи. У черепах нет зубов, но вместо них на клюве имеются твёрдые выпуклости и шероховатости. У хищных черепах они острые как ножи и служат для разрезания добычи. У травоядных черепах выпуклости снабжены зубчатыми краями, которые приспособлены для поедания твёрдой растительной пищи. Язык у черепах задействован при глотании, однако, в отличие от большей части рептилий, они не могут высовывать его наружу для захвата пищи.
Голова у большинства черепах относительно небольшая и имеет обтекаемую форму, чтобы при опасности легко скрываться в панцире. У мягкотелых черепах кончик морды вытянут в мягкий неподвижный хоботок, на конце которого размещаются ноздри :166 . Крупными, не умещающимися или плохо умещающимися под панцирь головами обладают черепахи, не полагающиеся на него как на основное средство защиты: морские черепахи , большеголовая черепаха , грифовая черепаха и так далее.







Шея черепах большинства видов умеренной длины или короткая, но у представителей рода змеиношейных черепах длина шеи почти равна длине карапакса :182 .
При испуге большинство черепах втягивает голову в панцирь, изгибая шею в форме S-образной кривой (при этом голова двигается прямо и назад). Однако существуют виды черепах (подотряд бокошейные черепахи (Pleurodira), распространённые в южном полушарии), которые прячут голову в панцирь, поворачивая её вбок в сторону плеча .

Строение конечностей черепах обусловлено их образом жизни. У сухопутных черепах задние конечности столбообразные, а передние несколько уплощённые, приспособленные для рытья нор. Большинство пресноводных черепах имеет плавательные перепонки между пальцами, а у морских черепах конечности преобразованы в ласты (задние короче передних) :75—76 .
Когда типичная черепаха прячется в своём панцире, передние конечности, защищённые спереди толстыми чешуями, прикрывают её голову.
Большинство черепах обладает коротким хвостом, который при втягивании в панцирь умещается между задними конечностями, прикрывая уязвимую область клоаки . У некоторых сухопутных черепах на кончике хвоста имеется шип заострённой ( балканская черепаха ) или ногтевидной ( плоскохвостая черепаха ) формы. Существуют черепахи с длинными хвостами. Как и в случае с крупной головой это виды, чей панцирь не несёт особой защитной функции: морские черепахи, каймановая черепаха , большеголовая черепаха и так далее.



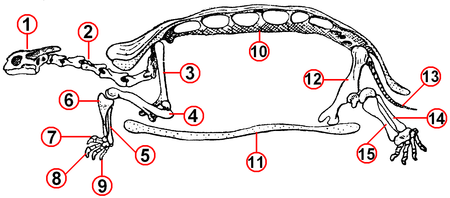
Череп панцирного типа — псевдостегальный (у современных морских черепах) или с ложной височной дугой — псевдоапсидный (у остальных черепах). Челюсти современных черепах лишены зубов и покрыты роговыми пластинками в виде клюва . Шейный и хвостовой отделы позвоночника подвижны, остальные срослись с карапаксом. Из-за развития панциря пояса конечностей у черепах переместились под рёбра (уникальное явление среди позвоночных) .

Панцирь — наиболее характерный видимый признак черепахи, отличающий её от других рептилий. Панцирь очень прочен: у некоторых видов он выдерживает тяжесть, в 200 раз превышающую массу самой черепахи [ источник не указан 483 дня ] . Чаще всего панцирь имеет два слоя: внутренний (костный) и наружный (кератиновый). Костный слой панциря составлен из пластинок, а роговой — из щитков. Высокая прочность панциря частично вызвана тем, что границы между внутренними пластинками и наружными щитками не совпадают .
Панцирь состоит из спинного щита — карапакса и брюшного — пластрона . Костная часть первого образована костными пластинками, сросшимися, как правило, с рёбрами и позвоночником . Второй у подавляющего большинства черепах образован грудиной , ключицами и брюшными рёбрами :113—114 . Сверху у большинства черепах панцирь покрыт симметричными роговыми щитками, и только у некоторых видов, таких как мягкотелые и кожистые черепахи, его покрывает плотная кожа :74 . Панцирь новорождённых черепах мягкий, но с возрастом он обычно твердеет :74—75 .

На спинном щите выделяются шейный, позвоночные, боковые (рёберные) и один ряд краевых щитков (у древнейшей известной науке черепахи с полностью сформированным панцирем Proganochelys quenstedti был двойной ряд краевых щитков), на брюшном — горловой, межгорловые, плечевые, грудные, брюшные, бедренные, заднепроходные, подмышечные, паховые и межкраевые щитки. Каждый щиток растёт самостоятельно, и если черепаха периодически впадает в спячку, то на нём вырастают концентрические годовые кольца .
Спереди и сзади панцирь имеет отверстия, через которые черепаха выдвигает голову, хвост и конечности. У некоторых видов подвижные части панциря могут плотно закрывать оба отверстия (или одно из них) при опасности. У коробчатых черепах передняя и задняя доли пластрона плотно притягиваются к карапаксу, надёжно защищая туловище со всех сторон . У паучьей черепахи ( Pyxis arachnoides ) подвижна только передняя доля пластрона. Специфическое строение имеет панцирь африканских киникс , у которых подвижна задняя доля не пластрона, а карапакса.
Окрас черепашьих панцирей бывает самым разным. Он может быть маскировочным (как у всех черепах России) или ярким и причудливым, как у лучистой и звёздчатой черепах . У многих видов на панцирях есть «украшения» в виде различных зубцов и гребней :75 .
Форма и вид панциря связана с образом жизни черепах: у сухопутных видов он чаще всего высокий, куполообразный, часто бугорчатый, у пресноводных — низкий, уплощённый и гладкий, у морских имеет обтекаемую каплевидную форму. Пластрон пресноводных и морских может быть частично редуцирован или расчленён на подвижные части, чтобы увеличить свободу движения. Чтобы компенсировать редукцию пластрона и уплощение карапакса, на последнем могут присутствовать упрочняющие его бугристые возвышения и «рёбра жёсткости» .

|

|
|
Карапакс черепахи
Роговые щитки: a — шейный; b — краевые; c — позвоночные; d — рёберные; e — надхвостовые |
Пластрон
зелёной черепахи
(Chelonia)
Роговые щитки: 1 — межгорловой; 2 — горловые; 3 — плечевые; 4 — грудные; 5 — брюшные; 6 — бедренные; 7 — анальные; 8 — подмышечные; 9 — нижнекраевые; 10 — паховые |
Наличие панциря тесно связано с особенностями строения внутренних органов, которые поэтому значительно отличаются от аналогичных органов других рептилий. Мускулатура туловища очень слабая, мышцы ног и шеи — наоборот, сравнительно сильны. Панцирь наделён осязанием — черепаха чувствует прикосновение к нему.
Рёберного дыхания у черепах нет; воздух заглатывается при помощи подъязычного аппарата либо движения конечностей. У мягкотелых черепах развито кожное дыхание. В воде черепахи дышат через кожу нёба, что позволяет не всплывать для дыхания во время зимовки. У некоторых видов пресноводных черепах газообмен отчасти происходит через специальные анальные мешки, открывающиеся в клоаку .
В отличие от большинства других рептилий, выделяющих во внешнюю среду в основном мочевую кислоту и часто вообще не имеющих мочевого пузыря , у черепах выделяется в основном мочевина и есть крупный мочевой пузырь .
Самцы водных черепах обычно имеют более длинный, чем у самок, хвост, а самцы сухопутных — хорошо выраженные . У тех и других на пластроне есть углубление, способствующее лучшей фиксации самца на самке при спаривании. Половой диморфизм проявляется также в размерах представителей разных полов, но здесь всё зависит от вида. Так, у звёздчатой черепахи ( Geochelone elegans ) самки крупнее самцов, а у шпороносной черепахи ( Centrochelys sulcata ) — наоборот. У некоторых видов самцы и самки различаются цветом радужной оболочки глаз . У самцов каролинской коробчатой черепахи глаза красного цвета, а у самок — жёлтого :161 .
Головной мозг у черепах развит слабо, его масса составляет всего 1/1000 долю массы тела. Спинной мозг , напротив, развит хорошо .
Черепахи обладают хорошо развитым цветовым зрением . При поиске пищи они в первую очередь ориентируются на её цвет и лишь затем на запах и вкус. Сухопутных черепах особенно сильно привлекает красный цвет. Они в первую очередь интересуются именно красными фруктами и овощами, а также совершенно несъедобными объектами этого цвета. Им нравится и зелёный цвет, но они предпочитают светло-зелёные оттенки тёмно-зелёным .
Уши черепах устроены довольно примитивно. У них нет ни ушных раковин, ни даже наружных слуховых проходов, а барабанная перепонка находится прямо на поверхности головы. Несмотря на анатомическую примитивность внешней части слухового аппарата у черепах хороший слух, хотя они воспринимают только низкие звуки частотой до 3000 Гц . У некоторых видов острота восприятия низких звуков сравнима с кошачьей .


В брачный период черепахи-самцы устраивают между собой турнирные бои за право спариться с самкой. У сухопутных черепах эти бои выражаются в преследовании соперника, попытках перевернуть его, нанесении ударов передним краем панциря и укусах, а у водных черепах это в основном укусы и преследование противника. Ухаживание победителя за самкой имеет цель заставить её остановиться и принять наиболее удобное для спаривания положение. Самцы расписной черепахи ( Chrysemys picta ) и украшенных черепах ( Pseudemys ) ухаживают за самкой оригинальным образом: они плывут задом наперед, увлекая за собой самку и поглаживая или похлопывая её морду длинными когтями своих передних лап. При спаривании пенис самца, обычно спрятанный у основания хвоста, выдвигается из клоаки . Спаривание у водяных черепах проходит в воде прямо во время плавания. При этом самец плотно фиксируется на самке, держа её клювом за кожу на шее :148—151 .
Самцы некоторых видов черепах в период спаривания могут «петь» — издавать несколько примитивных звуков .
Самки некоторых видов черепах способны подолгу сохранять в организме жизнеспособную сперму и делать несколько кладок без новых спариваний, но число яиц в кладке будет с каждым разом всё меньше .
Все известные современные черепахи — яйцекладущие. Самка может перекрыть доступ кислорода, чтобы «заморозить» развитие эмбриона, если условия среды неблагоприятны . Самки откладывают яйца в кувшинообразную ямку, которую они выкапывают задними лапами (некоторые — в собственные жилые норы ( гофер ) или в гнёзда крокодилов, или в морской прибрежный песок), иногда смачивая грунт жидкостью из клоаки :151—152 . Затем ямка засыпается и утрамбовывается сверху ударами пластрона. Яйца шарообразные или эллипсоидные, белого цвета, обычно покрыты твёрдой известковой скорлупой. Лишь у морских и некоторых бокошейных черепах яйца имеют мягкую кожистую оболочку. Число откладываемых яиц колеблется у разных видов от одного до двух сотен :26 .
Многие черепахи делают несколько кладок в течение сезона. Инкубационный период у большинства видов длится 2—3 месяца (у слоновой черепахи — 6—7 месяцев) . Пол морских черепах зависит от температуры инкубации яиц: если яйцо развивалось при более низкой температуре, то из яйца выводится самец, если при более высокой — самка .
У гигантских черепах ( Megalochelys gigantea ) отмечено наличие поведенческих механизмов регулирования численности популяции. При высокой плотности популяции самка откладывает всего 4—5 яиц раз в несколько лет. А при низкой плотности откладывается до 14 яиц несколько раз в год :151 .
Черепахи наиболее уязвимы на стадии яйца и новорождённого детёныша. Большая их часть гибнет именно в этот период. На затвердение панциря разным видам нужно разное время, но, в основном, несколько месяцев. Чем крупнее черепаха, тем меньше хищников ей угрожает, поэтому им надо быстро расти. Галапагосские черепахи в неволе, начиная примерно с 11 кг, ежегодно прибавляли столько же, пока их вес не достиг более 100 кг. Многие мелкие черепахи становятся половозрелыми в возрасте от 2 до 11 лет .
Черепахи большинства видов после откладывания яиц теряют к ним интерес и не проявляют никакой заботы о потомстве, но есть и исключения. Самки коричневой черепахи ( Manouria emys ) строят для своих яиц специальное гнездо и охраняют его вплоть до вылупления детёнышей .
Ещё один вид, проявляющий заботу о потомстве — ( Pseudemys malonei ). Наблюдали, как самка этого вида перед вылуплением отыскивала кладку и раскапывала её передними лапами, облегчая выход детёнышей :130 .
Черепахи ведут одиночный образ жизни и обычно ищут общества себе подобных только в брачный период, хотя некоторые виды могут собираться в группы на время зимовки. Некоторые пресноводные черепахи даже вне брачного периода агрессивно реагируют на своих сородичей, например жабоголовые черепахи ( Phrynops geoffroanus ) . Красноухие черепахи ( Trachemys scripta ) по очереди очищают панцири друг друга от водорослей :29 .
Сухопутные черепахи питаются в основном растительной пищей, а пресноводные, наоборот, в большинстве своём хищники. Они поедают различных рыб , а также моллюсков , членистоногих и других беспозвоночных . Но и те и другие охотно включают в свой рацион корм, казалось бы, им несвойственный: растительноядные — животную пищу, хищники — растительную. Среди морских черепах есть и хищные, и растительноядные ( зелёная черепаха ) , и всеядные виды :80 . Многие пресноводные виды черепах в детстве ведут хищнический образ жизни, а повзрослев, становятся растительноядными.

Черепахи — рекордсмены по долголетию среди всех позвоночных животных. Гигантская черепаха по имени Айвата прожила не менее 150, а возможно, и 250 лет , а другой представитель этого вида по кличке Кики — не меньше 146 лет . Слоновая черепаха Гариетта дожила до 175 лет . Мадагаскарская лучистая черепаха по имени Туи Малила умерла в возрасте не менее 188 лет .
Ядовитость — редкое явление среди черепах. Мясо некоторых видов черепах может быть ядовитым, так как рацион этих видов включает в себя токсичные организмы. Среди сухопутных черепах ядовитым может быть мясо коробчатых черепах , в рацион питания которых входят ядовитые грибы , а среди морских черепах известны случаи отравления мясом черепах бисс и кожистых черепах , которые поедают ядовитых медуз .

Разнообразные виды черепах населяют бо́льшую часть суши и вод тропического и умеренного поясов от 56° с. ш. (Европейская болотная черепаха в Латвии) до 42 ю. ш. ( Аргентинская черепаха в Аргентине) . Их нет в холодных краях, некоторых пустынях, на тихоокеанском побережье Южной Америки , и в Новой Зеландии .
На территории России встречаются :
В Казахстане и странах Средней Азии обитает также среднеазиатская черепаха ( Agrionemys horsfieldii ) .
В красной книге Международного союза охраны природы ( МСОП ) внесены данные об охранном статусе 228 из 328 современных видов черепах. Из них 6 считаются вымершими, а ещё 135 находятся под угрозой вымирания (классификации МСОП: EW, CR, EN, VU). Среди основных факторов человеческого вмешательства увеличивающих угрозу выживанию различных видов черепах МСОП отмечает отлов черепах (угрожает 92 видам), загрязнение среды обитания (41 вид) и создание транспортных сетей (41 вид). Дополнительными факторами угрозы служит сокращение среды обитания черепах вследствие развития сельского хозяйства и жилых районов. На первом месте среди естественных факторов угрозы для черепах стоит угроза со стороны других видов животных (местных и инвазивных) .

Черепашье мясо — основная причина промысла многих видов черепах человеком (за исключением самых мелких) .
В конце XX века только у берегов Сомали ( Восточная Африка ) рыбаки ежегодно вылавливали до 4000 зелёных черепах. Наряду с зелёной черепахой сильно пострадала и атлантическая ридлея ( Lepidochelys kempii ). С середины 50-х годов до середины 70-х годов XX века её численность катастрофически сократилась и в 1982 году сохранилось не более нескольких сотен этих животных. Промысел морских черепах и по сей день распространён в Карибском море, причём, по данным на конец XX века, местные рыбаки теперь добывают только идущую на изготовление деликатесного супа хрящевую ткань , так называемый «калиппи». От черепахи весом 120 кг можно получить всего лишь около 2,5 кг «калиппи». :8—10 .

Морских черепах ловят в основном на берегу, куда самки выползают для кладки яиц. В воде этих животных ловят при помощи сетки с кольцом. Кольцо набрасывают на черепаху и она, испугавшись, сама запутывается в сетке. В некоторых районах применяют старинный способ ловли черепах с помощью рыбы-прилипалы . Жители Мозамбика и Мадагаскара привязывают к хвосту пойманной прилипалы верёвку и бросают в море неподалёку от черепахи. Рыба немедленно крепко присасывается к черепахе, и остаётся только вытащить их обеих из воды . «Грузоподъёмность» одной рыбы составляет около 30 кг, поэтому для охоты на черепах применяют обычно сразу нескольких прилипал на одном лине. Вместе они могут удержать черепаху в несколько центнеров весом :43 .
Ряд видов и подвидов гигантских сухопутных черепах, обитавших на тропических островах, был истреблён вследствие чрезмерного промысла моряками. Согласно записям в судовых журналах, только 79 китобойных судов за 36 лет в середине XIX века вывезли с Галапагосских островов 10 373 слоновые черепахи ( Chelonoidis elephantopus ). Всего за XVII — XVIII века , по данным архивных материалов, на Галапагосах было уничтожено около 10 миллионов этих животных, причём на островах и они были истреблены, а на ряде других оказались на грани исчезновения. Огромное количество гигантских черепах ( Megalochelys gigantea ) было также вывезено с Сейшельских , Маскаренских и других островов Индийского океана , на некоторых из которых они полностью исчезли. Есть сведения, что в начале XVIII века на острове Реюньон на корм свиньям было пущено около 40 000 особей одного из обитавших там и исчезнувших ныне видов :8—10 . Использование сухопутных черепах на корм животным практиковалось и в СССР — там ежегодно заготавливалось более 100 000 среднеазиатских черепах , которые шли в основном на откорм пушных зверей :230 .
Разнообразием отличаются методы, применяемые современными охотниками за черепахами. Их добывают острогой, ловят сетями, ловушками или на крючок. Пресноводных черепах выкапывают из-под ила, когда водоёмы пересыхают в сухой сезон и черепахи впадают в спячку. На лесных черепах (например, Cuora galbinifrons , Heosemys spinosa , Indotestudo elongata или индийская шиповатая черепаха ) охотятся с собаками .
Многие популяции сухопутных черепах сокращаются вследствие застройки или использования в других хозяйственных целях населяемых ими земель . В некоторых случаях землепользователи делают попытки переселить охраняемых законом черепах с осваиваемых земель на другие территории, но такие программы не всегда успешны. К примеру, в марте 2008 года армия США перевезла вертолётами 670 пустынных западных гоферов ( Gopherus agassizii ) с территорий, на которых создаётся новый ( англ. ), в другие части пустыни Мохаве . Однако, как было вскоре обнаружено, перемещённые черепахи плохо приживаются на новом месте и часто поедаются местными койотами . Это привело к приостановке программы переселения .

Мясо черепах употребляется в пищу как в сыром виде, так и в варёном ( суп ) или жареном ( жаркое ).
Неприхотливость черепах, облегчающая их транспортировку в живом виде, привела к тому, что моряки стали использовать черепах как «живые консервы»
Начало применения черепахового панциря, как облицовочного материала и для декорирования различных, в том числе и ювелирных изделий, а также для инкрустаций, относится к 80 г до н. э. Черепаший панцирь использовался как материал для подделки дерева, украшения предметов интерьера листами (пластинами) поверх отдельного элемента. Продавались роговые пластины, покрывающие костистый верхний слой черепахи. Цена таких пластин зависела от уровня их прозрачности, интенсивности окраски, а также способности к их полировке.
При термообработке, черепаховый щит размягчается при температуре кипения воды (100 °C). При высоких температурах, щит темнеет, а его полупрозрачность уменьшается, поэтому пластины отделяют от верхнего щита, при самых низких температурах. Полученные изогнутые пластины выравнивают с помощью пресса . Для увеличения площади пластинок, правильно скошенные края накладывают друг на друга и спаивают вместе щипцами или в тисках, равномерно нагревая кипящей водой или подвергая равноценному сухому обогреву.
В наши дни черепаший панцирь используется для декорирования мелких деталей мебели (ручек, замков, выступающих узоров), в украшениях (заколки, серьги, браслеты) и так же для: гребней для волос, ручек для ножей и т. д. [ источник не указан 1703 дня ]
Из однородного черепахового панциря вырезаются японские традиционные женские украшения для волос — кандзаси .
С целью изучения особенностей образа жизни и поведения слоновых черепах, обитающих на Галапагосских островах , в мае 2009 года учёные прикрепили к нескольким черепахам видеокамеры (« Криттеркам ») .
В 1968 году советский космический аппарат « Зонд-5 » впервые в мире без пилотов облетел вокруг Луны . На его борту находились две среднеазиатские черепахи . Они стали первыми живыми существами в истории, возвратившимися на Землю после облёта Луны — за три месяца до полёта « Аполлона-8 » .

Некрупные сухопутные и пресноводные черепахи различных видов являются популярными домашними питомцами. В Европе наибольшей популярностью пользуются такие сухопутные черепахи, как средиземноморская , балканская и среднеазиатская . Их продажа составила в 1938 году 250 000 особей и долгое время держалась на уровне 100 000 экземпляров в год. Они завозились из Марокко , Турции , Испании , Греции , Болгарии , а также из СССР, откуда с 1967 года из Казахстана и бывших республик Средней Азии на мировой рынок поставлялось около 1 миллиона среднеазиатских черепах. В 1971 году одна только Югославия экспортировала в другие страны Европы (в основном ФРГ , Италию и Голландию ) около 40 000 балканских черепах. С 1969 по 1973 год в Англию было завезено почти 1,5 миллиона черепах различных наземных видов, а ещё 41 694 особи пресноводных видов европейская болотная черепаха ( Emys orbicularis ) и Mauremys caspica . объём мировых продаж черепах для содержания в неволе составил приблизительно 10 миллионов мелких черепах :8—10 .
В России из четырёх видов наземных черепах, обитающих сегодня на её территории, в неволе чаще всего содержатся два: болотная ( Emys orbicularis ), населяющая низовья Волги и в другие южные регионы европейской части России, и средиземноморская ( Testudo graeca ), распространённая на черноморском побережье Кавказа и в Дагестане . Эти виды занесены в Международную Красную книгу , а средиземноморская — также и в Красную книгу РФ в категории «1» (вид с неуклонно сокращающейся численностью, отдельные популяции которого находятся на грани исчезновения) и в приложение к Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения.
Зарегистрированы случаи контрабанды черепах в Россию .
В США наиболее распространёнными домашними черепахами являются красноухая черепаха и расписная черепаха . По состоянию на 2010 год, большинство американских штатов разрешают, но не рекомендуют заводить черепах в качестве домашних животных. В Орегоне их содержание дома запрещено , а в Индиане запрещена торговля ими .
Федеральный закон США запрещает торговлю или транспортировку черепах размером менее 10 см, чтобы оградить людей от контакта с переносчиками сальмонеллы . Тем не менее это разрешено в исследовательских целях; также известны случаи нелегальной перевозки маленьких черепах .
На Среднем Западе США черепашьи бега являются популярным развлечением во время ярмарок .
В отличие от змей , черепахи практически не представляют угрозы для людей. Как исключения — самец кожистой черепахи в брачный период может принять пловца за самку, схватить его ластами и утопить , а кусачая и агрессивная каймановая черепаха способна по ошибке или обороняясь серьёзно укусить человека .
|
|
В этом разделе
не должны перечисляться малозначимые либо слабо связанные с объектом статьи его упоминания и изображения
в произведениях культуры
.
|


Черепахи занимают заметное место в мифологии и культуре многих народов . Особенно значительную роль они играли в мифологии Китая , где черепаха , наряду с драконом , цилинем и фениксом , традиционно причислялась к « четырём благопожелательным существам » — а в доисторические времена, черепаха, возможно, считалась и моделью мироздания .
В мифологии народов Африки Земля изображается в образе черепахи, поскольку её внешний облик и окрас придают ей сходство с почвой . Черепаха также является популярным персонажем африканских сказок , ей приписываются сверхъестественные способности и причастность к магии .
Форма черепахи . Каменные черепахи уже более полутора тысяч лет гордо несут стелы китайских императоров . Они украшают фонтаны в Италии и Испании.
А 21 мая 2009 год и в латвийском городе Даугавпилс был торжественно открыт европейской болотной черепахе .
Во время Второй мировой войны черепаха служила символом саботажа в борьбе польского Сопротивления (лозунг « работай медленно »).
Черепаха Квази — персонаж « Алисы в Стране чудес » Льюиса Кэрролла .
Черепашки-ниндзя — вымышленная команда четырёх антропоморфных черепах-мутантов.
Великий А’Туин — вымышленная гигантская черепаха, на спине которой происходит действие в Плоском мире Терри Пратчетта .
Лев-черепаха — персонаж мультсериала « Аватар: Легенда об Аанге », обучивший Аанга магии энергии.
Торатон — гигантская (размером с крупного динозавра- завропода ) черепаха во 2-й серии фильма « Дикий мир будущего ».
Сквиртл — персонаж франшизы « Покемон », выглядит как синяя черепаха с красным панцирем.
Широко известен « парадокс » Зенона — « Ахиллес и черепаха ». Это одна из апорий Зенона : « Быстроногий Ахиллес никогда не догонит черепаху, если в начале движения черепаха находится впереди на некотором расстоянии от него, а он с каждым шагом сокращает расстояние в два раза».
Аристотель в своей «Физике» подробно анализирует рассуждения Зенона. Этот сюжет многократно использовался в поэзии и сатире .



![Герб деревни Хёнов[de] в Германии](/images/004/972/4972156/65.jpg?rand=827159)


























(2021) рекомендует следующую классификацию современных черепах до семейств включительно :
Ранее была распространена классификация с выделением 5 современных и 2 вымерших подотрядов:
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
Захваченных черепах обыкновенно переворачивают на спину. С ними не церемонятся: их просто складывают в сторонке где-нибудь на палубе, протягивают над ними парус для защиты от солнца и не заботятся более ни о чём, полагаясь на их живучесть. Ни пищи, ни питья им не дают… В таком положении черепахи могут совершать продолжительные путешествия.
{{
cite AV media
}}
:
Проверьте значение даты:
|accessdate=
(
справка
)
. Filmmaker
, the creator of the film
, talks to
, the director of conservation genetics at the
American Museum of Natural History
about turtle conservation and the relationship between evolution and extinction
{{
cite news
}}
:
Проверьте значение даты:
|accessdate=
(
справка
)
;
Недопустимый
|ref=harv
(
справка
)
;
Неизвестный параметр
|month=
игнорируется (
справка
)