Бешуля, Спиридон Ерофеевич
- 1 year ago
- 0
- 0

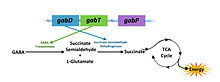
ГАМК-оперон несет ответственность за превращение γ-аминобутирата ( ГАМК ) в сукцинат . ГАМК - оперон включает три структурных гена - gabD , gabT и gabP , которые кодируют сукцинат полуальдегида дегидрогеназы, ГАМК-трансаминазы и ГАМК-пермеазы соответственно. Существует регулирующий ген csiR , ниже оперона, который кодирует предполагаемый репрессор транскрипции и активируется при ограничении азота.
ГАМК-оперон был выявлен в кишечной палочке и значимые гомологи для ферментов были обнаружены в организмах, таких как Saccharomyces cerevisiae , крысы и человека .
Ограничение азота — условие активации генов ГАМК. Ферменты, полученные этими генами преобразуют ГАМК в сукцинат, который затем входит в ЦТК, чтобы использоваться в качестве источника энергии. ГАМК-оперон также известен содействием гомеостазу полиаминов в течение роста при ограничении азота и поддержанию высоких концентраций внутреннего глутамата в условиях стресса.
ГАМК-оперон состоит из трех структурных генов:
Ген GabT кодирует ГАМК-трансаминазу , фермент , который катализирует превращение ГАМК и 2-оксоглутарат в полуальдегид сукцината и глутамата. Полуальдегид сукцината затем окислятся в сукцинат с помощью полуальдегида дегидрогеназы сукцината (который кодируется геном gabP), тем самым вводя в ЦТК в качестве полезного источника энергии. ГАМК-оперон способствует гомеостазу полиаминов, таких как путресцин в ходе роста при ограниченнии азота. Известно также о его роли в поддержании высоких концентраций внутреннего глутамата в условиях стресса.
Экспрессия генов в опероне управляется тремя дифференциально регулируемыми промоторами , два из которых управляются , кодирующим сигма-фактор σ S .
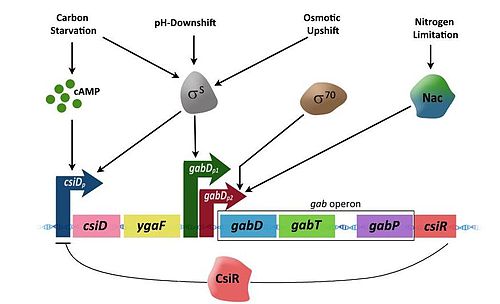
Промотор csiD ( csiD p ) имеет важное значение для экспрессии csiD (генов, индуцированных углеродным голоданием), ygaF и ГАМК-генов. СsiD p активирован только в условиях углеродного голодания и стационарной фазы, в течение которой цАМФ накапливается в высоких концентрациях в клетке. Связывание цАМФ с белком рецептора цАМФ (CRP) вынуждает CRP жестко связываться с определенным сайтом ДНК в промоторе 'csiD p , таким образом, активируя транскрипцию генов ниже промотора.
gabD p1 оказывает дополнительное управление gabDTP в регионе. gabD p1 активирует σ S вызывая условия, такие как гиперосмотические и кислые сдвиги, кроме голодания и стационарной фазы. Промотор gabD p2 , с другой стороны, является σ 70 -зависимым и активируется при ограничении азота. В условиях ограничения азота, регулятор азота Nac связывается с участком, расположенного чуть выше промотора, экспрессирующего гены ГАМК. Гены ГАМК при активации вырабатывают ферменты, которые превращают ГАМК в сукцинат.
Наличие азота активирует ген csiR , расположенный ниже гена gabP . Ген csiR кодирует белок, который действует как репрессор транскрипции для csiD-ygaF-ГАМК оперонов, следовательно, отключая деградацию пути ГАМК.
Деградации путей ГАМК существует почти во всех эукариотических организмах и происходит под действием аналогичных ферментов. Хотя, ГАМК в кишечной палочке в основном используется в качестве альтернативного источника энергии, ГАМК в высших эукариотических организмах выступают в качестве тормозящего нейромедиатора , а также в качестве регулятора мышечного тонуса. Пути деградации ГАМК у эукариот несут ответственность за инактивацию ГАМК.