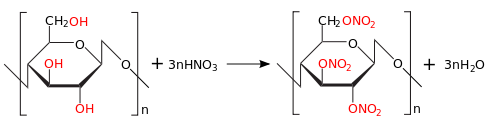
Нитроцеллюлоза
- 1 year ago
- 0
- 0
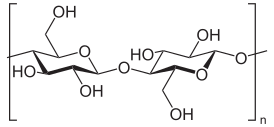
Целлюло́за , клетчáтка ( фр. cellulose от лат. cellula — «клетка») — органическое соединение , углевод , полисахарид с формулой (C 6 H 10 O 5 ) n . Молекулы — неразветвлённые цепочки из остатков β- глюкозы , соединённых гликозидными связями β-(1→4). Белое твёрдое вещество, нерастворимое в воде. Главная составная часть клеточных оболочек всех высших растений .
Целлюлоза была обнаружена и описана французским химиком Ансельмом Пайеном в 1838 году.
Целлюлоза представляет собой линейный гомополимер из сотен или десятков тысяч остатков D-глюкозы . Соединение фрагментов глюкозы обеспечивается β(1→4)- гликозидной связью . Такое соединение мономерных звеньев отличает целлюлозу от α(1→4)-гликозидных связей характерных для других гомополимеров глюкозы: крахмала и гликогена . В отличие от амилозы крахмала, молекулы которой сворачиваются в спираль, макромолекула целлюлозы склонна принимать вытянутую стержневую конформацию.

Целлюлоза — белое твёрдое, стойкое вещество, не разрушается при нагревании (до 200 °C). Является горючим веществом, температура разложения — 275 °С , температура самовоспламенения — 420 °С (хлопковая целлюлоза). В 2016 году экспериментально показано плавление целлюлозы при 467 °C .
Нерастворима в воде, слабых кислотах и большинстве органических растворителей. Однако благодаря большому числу гидроксильных групп является гидрофильной ( краевой угол смачивания составляет 20—30 градусов) .
Целлюлоза не имеет вкуса и запаха. Зарегистрирована в качестве пищевой добавки E460.
Целлюлоза подвергается биодеградации при участии многих микроорганизмов .
Целлюлоза состоит из остатков молекул глюкозы , которая и образуется при гидролизе целлюлозы:
Серная кислота с иодом , благодаря гидролизу , окрашивают целлюлозу в синий цвет.
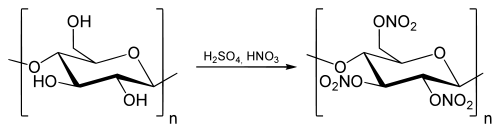
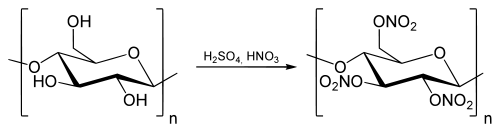
При реакции с азотной кислотой образуется нитроцеллюлоза ( тринитрат целлюлозы ):
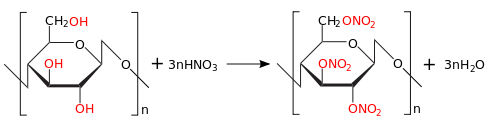
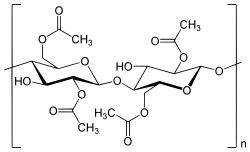
В процессе этерификации целлюлозы уксусной кислотой получается триацетат целлюлозы :

Целлюлозу крайне сложно растворить и подвергнуть дальнейшим химическим превращениям, однако в среде подходящего растворителя, например, в ионной жидкости , такой процесс можно осуществить эффективно .



Промышленным методом целлюлозу получают методом варки щепы на целлюлозных заводах, входящих в промышленные комплексы ( комбинаты ). По типу применяемых реагентов различают следующие способы варки целлюлозы:
Получаемая после варки техническая целлюлоза содержит различные примеси: лигнин , гемицеллюлозы . Если целлюлоза предназначена для химической переработки (например, для получения искусственных волокон), то она подвергается облагораживанию — обработке холодным или горячим раствором щелочи для удаления гемицеллюлоз .
Для удаления остаточного лигнина и придания целлюлозе белизны проводится её отбелка. Традиционная для XX века хлорная отбелка включала в себя две ступени:
С 1970-х годов в практику вошла также отбелка озоном. В начале 1980-х годов появились сведения об образовании в процессе хлорной отбелки чрезвычайно опасных веществ — диоксинов . Это привело к необходимости замены хлора на другие реагенты. В настоящее время технологии отбелки подразделяются на:
Используется в качестве наполнителя в таблетках в фармацевтике. Целлюлозу и её эфиры используют для получения искусственного волокна ( вискозного , ацетатного , медно-аммиачного шёлка , искусственного меха ). Хлопок , состоящий большей частью из целлюлозы (до 99,5 %), идёт на изготовление тканей .
Древесная целлюлоза используется для производства бумаги , пластмасс , кино- и фотоплёнок , лаков , бездымного пороха и т. д.
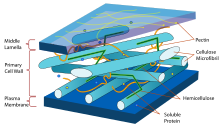
Целлюлоза является одним из основных компонентов клеточных стенок растений, хотя её содержание в различных клетках или даже частях стенки одной клетки сильно варьируется. Так, например, стенки клеток эндосперма злаков содержат всего около 2 % целлюлозы, в то же время хлопковые волокна , окружающие семена хлопчатника , состоят из целлюлозы более чем на 90 %. Клеточные стенки в области кончика удлинённых клеток, характеризующихся полярным ростом ( пыльцевая трубка , корневой волосок ), практически не содержат целлюлозы и состоят в основном из пектинов , в то время как базальные части этих клеток содержат значительные количества целлюлозы. Кроме того, содержание целлюлозы в клеточной стенке изменяется в ходе онтогенеза , обычно вторичные клеточные стенки содержат больше целлюлозы, чем первичные.
Отдельные макромолекулы целлюлозы включают от 2 до 25 тысяч остатков D-глюкозы. Целлюлоза в клеточных стенках организована в микрофибриллы, представляющие собой паракристаллические ансамбли из нескольких отдельных макромолекул (у сосудистых растений около 36) , связанных между собой водородными связями и силами Ван-дер-Ваальса .
Макромолекулы, находящиеся в одной плоскости и связанные между собой водородными связями, формируют лист в пределах микрофибриллы. Между собой листы макромолекул также связаны большим числом водородных связей. Хотя водородные связи довольно слабые, благодаря их большому количеству микрофибриллы целлюлозы обладают высокой механической прочностью и устойчивостью к действию ферментов .
Индивидуальные макромолекулы в микрофибрилле начинаются и заканчиваются в разных местах, поэтому длина микрофибриллы превышает длину отдельных макромолекул целлюлозы. Макромолекулы в микрофибрилле ориентированы одинаково, то есть редуцирующие концы (концы со свободной, аномерной OH-группой при атоме C 1 ) расположены с одной стороны.
Современные модели организации микрофибрилл целлюлозы предполагают, что в центральной области она имеет высокоорганизованную структуру, а к периферии расположение макромолекул становится более хаотичным. Так, в центре микрофибриллы высших растений располагается ядро из 24 молекул. Ещё 12 молекул расположены по периферии фибриллы. Теоретически диаметр такой микрофибриллы составляет 3,8 нм, однако, данные рентгеноструктурного анализа показывают, что это значение несколько меньше — 3,3 нм, что соответствуют 24 молекулам. По-другим оценкам размеры фибрилл значительно больше: 5—9 нм в поперечном сечении (более 50 отдельных макромолекул).
Между собой микрофибриллы связаны сшивочными гликанами ( гемицеллюлозы ) и, в меньшей степени, пектинами . Целлюлозные микрофибриллы, связанные сшивочными гликанами, формируют трёхмерную сеть, погружённую в гелеобразный матрикс из пектинов и обеспечивающую высокую прочность клеточных стенок.
Во вторичных клеточных стенках микрофибриллы могут быть ассоциированы в пучки, которые называют макрофибриллами. Подобная организация дополнительно увеличивает прочность клеточной стенки.
Образование макромолекул целлюлозы клеточных стенок высших растений катализирует мультисубъединичный мембранный целлюлозосинтазный комплекс, расположенный на конце удлиняющихся микрофибрилл. Полный комплекс целлюлозосинтазы состоит из каталитической, поровой и кристаллизационной субъединиц. Каталитическая субъединица целлюлозосинтазы кодируется мультигенным семейством CesA ( cellulose synthase A ), которое входит в суперсемейство Csl ( cellulose synthase-like ), включающее также гены CslA , CslF , CslH и CslC , ответственные за синтез других полисахаридов.
При изучении поверхности плазмалеммы растительных клеток методом замораживания-скалывания в основании целлюлозных микрофибрилл можно наблюдать так называемые розетки или терминальные комплексы размером около 30 нм и состоящие из 6 субъединиц. Каждая такая субъединица розетки является в свою очередь суперкомплексом, образованным из 6 целлюлозосинтаз. Таким образом, в результате работы подобной розетки формируется микрофибрилла, содержащая на поперечном срезе около 36 макромолекул целлюлозы. У некоторых водорослей суперкомплексы синтеза целлюлозы организованы линейно.
Интересно, что роль затравки для начала синтеза целлюлозы играет гликозилированный ситостерин . Непосредственным субстратом для синтеза целлюлозы является UDP-глюкоза. За образование UDP-глюкозы отвечает сахарозосинтаза, ассоциированная с целлюлозосинтазой и осуществляющая реакцию:
Кроме того, UDP-глюкоза, может образовываться из пула гексозофосфатов в результате работы УДФ-глюкозопирофосфорилазы:
Направление синтеза микрофибрилл целлюлозы обеспечивается за счёт движения целлюлозосинтазных комплексов по микротрубочкам , прилежащим со внутренней стороны к плазмалемме. У модельного растения резуховидка Таля обнаружен белок CSI1, отвечающий за закрепление и движение целлюлозосинтазных комплексов по кортикальным микротрубочкам.
У млекопитающих (как и большинства других животных) нет ферментов , способных расщеплять целлюлозу. Однако многие травоядные животные (например, жвачные ) имеют в пищеварительном тракте бактерии-симбионты, которые расщепляют и помогают хозяевам усваивать этот полисахарид. Расщепление целлюлозы связано с действием в расщепляющих организмах фермента целлюлазы . Бактерии, расщепляющие целлюлозу, называемые целлюлозоразрушающими ( англ. cellulolytic bacteria ), это часто актинобактерии рода Cellulomonas , являющиеся факультативными анаэробами , аэробные бактерии рода Cellvibrio . Однако, например, для бумажных книг они представляют опасность только при их намокании, когда кожа и клей начинают разрушаться гнилостными бактериями, а бумага и ткани — целлюлозоразрушающими . Очень опасны для бумажных книг плесневые грибы , разрушающие целлюлозу. За три месяца они могут разрушить 10—60 % волокон бумаги, благоприятные условия для их развития — влага и воздух повышенной влажности , наиболее благоприятная температура — от +22 до +27 градусов Цельсия, они могут распространяться от поражённых ими книг на другие . Активно расщепляющие целлюлозу плесневые грибы — это, например, Chaetomium globosum , .
|
Мультисахариды
|
|
|---|---|
| Дисахариды | |
| Трисахариды | |
| Тетрасахариды | |
|
|
|
|
|
|
| Олигосахариды | |
| Полисахариды | |
| Органеллы |
|
||||
|---|---|---|---|---|---|
| Клеточная стенка |
|
||||
|
Деление
растительной клетки |
|
||||