Динозавры Северной Америки
- 1 year ago
- 0
- 0

Диноза́вры ( лат. Dinosauria ) — группа архозавров из клады авеметатарзалий . Динозавры возникли в триасовом периоде , между 243 млн и 233,23 млн лет назад, и стали доминирующими наземными позвоночными после триасово-юрского вымирания 201 млн лет назад; их доминирование продолжалось на протяжении остальной части мезозойской эры , в течение юрского и мелового периодов. Хотя традиционно динозавры рассматривались как полностью вымершая группа в ранге надотряда , большинство современных специалистов придерживается принципов кладистической классификации , в соответствии с которыми птицы являются ныне живущими оперёнными динозаврами из клады теропод , возникшими во время верхнеюрской эпохи , либо в меловом периоде (в зависимости от определения таксона ), и, в отличие от всех остальных динозавров, пережившими мел-палеогеновое вымирание 66 млн лет назад. Условно всех динозавров, за исключением птиц, называют нептичьими динозаврами .
Динозавры — разнообразная группа с таксономической , морфологической и экологической точек зрения. Используя ископаемые свидетельства, палеонтологи выявили более 900 различных родов и более 1000 видов нептичьих динозавров. Динозавры представлены на каждом континенте , как современными видами (птицами), так и ископаемыми. Птицы, насчитывающие более 10 800 современных видов, являются одной из самых разнообразных групп позвоночных . , показала, что нептичьи динозавры, как и птицы, были активными теплокровными животными с повышенным обменом веществ и многочисленными приспособлениями к социальным взаимодействиям. Динозавры заняли многочисленные экологические ниши и освоили разнообразные типы питания. Все динозавры — яйцекладущие животные, причём способность к строительству гнёзд разделяется как птицами, так и многими нептичьими динозаврами.
Хотя предки динозавров передвигались на двух конечностях , многие нептичьи динозавры ходили на четырёх , а некоторые из них были способны менять эти позиции. Сложные демонстрационные структуры, такие как рога и гребни, присутствовали у всех групп динозавров, а некоторые их вымершие представители развили такие модификации, как костяная броня и шипы. В то время как современные птицы, как правило, невелики из-за ограничений размера, необходимых для полёта, многие доисторические динозавры, в том числе и птицы, достигали больших размеров — рекордсменами в этом плане являлись завроподы группы титанозавров , среди которых были крупнейшие когда-либо существовавшие наземные животные. Тем не менее, идея о том, что все нептичьи динозавры обладали гигантскими размерами, является ошибочным представлением, основанным частично на том, что вероятность разрушения больших и прочных костей до того, как они окаменеют, гораздо меньше, чем вероятность того, что то же самое произойдёт с костями маленького животного.
С тех пор, как первые окаменелые кости динозавров были опознаны и признаны в начале XIX века, их ископаемые скелеты стали главными достопримечательностями музеев по всему миру, а динозавры превратились в неотъемлемую часть популярной культуры . Большие размеры представителей некоторых групп динозавров, а также распространившийся чудовищный и фантастический образ, обеспечили регулярное появление динозавров в бестселлерах и фильмах, таких как « Парк юрского периода ». Постоянный общественный интерес к этим животным привёл к значительному финансированию палеонтологии, и новые открытия регулярно освещаются в средствах массовой информации .
Термин «динозавр» ввёл в 1842 году английский биолог Ричард Оуэн для описания первых ископаемых остатков древних ящеров, поражавших воображение учёных своими размерами . Слово происходит от древнегреческого — «страшный» и — «ящер». Хотя таксономическое имя часто интерпретировалось как некое указание на зубы, когти и другие внушающие страх особенности динозавров, на самом же деле Оуэн дал этот термин, указывая на их размер и величие . Поясняя свои взгляды, Оуэн называл динозавров толстокожими вторичного периода . К толстокожим в XIX веке относили крупных и массивнотелых бегемотов , слонов и носорогов , а вторичным периодом считали мезозойскую эру.

Динозавры относятся к группе архозавров , которая подразделяется на псевдозухий , то есть крокодилов и их родственников, и авеметатарзалий , то есть птиц и их родственников, таких как нептичьи динозавры и птерозавры .
В динозавры обычно определяются как группа, состоящая из ближайшего общего предка (БОП) трицератопса и современных птиц , а также всех его потомков . Кроме того, существует другой вариант определения, согласно которому к динозаврам относится БОП мегалозавра и игуанодона и все его потомки; оно связано с тем, что мегалозавр и игуанодон — два из трёх родов, первыми включённых в состав динозавров .
Оба этих определения подразумевают разделение клады динозавров на две основные ветви: птицетазовые , или орнитисхии (Ornithischia), и ящеротазовые , или заврисхии (Saurischia) . К птицетазовым относят таксоны, обладающие более поздним общим предком с трицератопсом, чем с тираннозавром , в то время как ящеротазовые, по традиционным представлениям о филогении динозавров, включают таксоны, обладающие более недавним общим предком с тираннозавром, чем с трицератопсом . Анатомически эти две группы наиболее заметно различаются в строении таза, хотя их названия носят несколько условный характер, так как птицы относятся не к птицетазовым, а к ящеротазовым, поскольку произошли от более ранних представителей этой группы . Эта путаница имеет историческую основу: в 1887 году, когда палеонтолог Гарри Сили предложил подразделять динозавров на птицетазовых и ящеротазовых, происхождение птиц от динозавров и, тем более, конкретно от ящеротазовых, было не более чем спорной гипотезой, а о включении их в состав этого таксона тогда не могло идти и речи .
У птицетазовых таз внешне напоминал птичий: их лобковая кость была ориентирована каудально , то есть направлена назад. В отличие от птиц, у птицетазовых лобковая кость обычно имела дополнительный отросток, направленный вперёд. В состав птицетазовых включают множество таксонов, представители которых в основном были растительноядными, среди которых можно выделить следующие наиболее разнообразные группы :
Ранние ящеротазовые сохранили структуру бедра своих предков с лобковой костью, направленной краниально или вперёд . Эта основная форма была модифицирована поворотом лобковой кости назад в различной степени в нескольких группах ( герреразавр , теризинозавроиды , дромеозавриды и птицы ). К ящеротазовым относят две группы :

|

|

|

|
| Тазовые кости птицетазовых. Лобковые кости направлены назад | Тазовые кости эдмонтозавра , представителя птицетазовых | Тазовые кости типичных ящеротазовых. Лобковые кости выступают вперёд | Тазовые кости тираннозавра , представителя ящеротазовых |

В соответствии со всеми существующими филогенетическими определениями, птицы являются ныне живущей группой динозавров-теропод. Хотя в традиционной систематике птицы считались отдельным классом , произошедшим от динозавров, отдельного надотряда , подавляющая часть современных палеонтологов, изучающих динозавров, отвергает традиционную систематику в пользу филогенетической таксономии. Этот подход предполагает, что для того, чтобы группа была естественной, все потомки членов группы также должны быть включены в неё. Таким образом, птицы в настоящее время, как правило, признаются ныне живущей группой динозавров . По мере обнаружения большего количества ископаемых теропод, имеющих большее или меньшее родство с современными птицами, граница между «птицами» и «не-птицами» становится всё более размытой. Птицы классифицируются как принадлежащие к подгруппе манирапторов , которая относится к целурозаврам , в свою очередь входящим в группу теропод .
Ниже представлена упрощённая кладограмма , отображающая традиционные представления о филогении динозавров :
| Динозавры |
|
||||||||||||||||||||||||||||||||||||||||||||||||
Обратите внимание, что прозавроподы и нептичьи тероподы не образуют реальные эволюционные ветви (клады), а состоят из множества отдельных ветвей, и поэтому отмечены здесь только для упрощения .

Согласно недавно выдвинутой альтернативной и довольно спорной точке зрения, завроподоморфы являются сестринской по отношению к остальным динозаврам группой, не попадающей под указанные выше определения. Для обеспечения таксономической стабильности было дано несколько новых «безопасных» определений, по которым к динозаврам обязательно относятся птицетазовые, завроподоморфы и тероподы. Например, М. Барон и коллеги (2017) определили Dinosauria как БОП трицератопса грубого ( Triceratops horridus ), домового воробья ( Passer domesticus ) и диплодока Карнеги ( Diplodocus carnegii ) и все его потомки, чтобы гарантировать, что завроподы и их родственники будут однозначно подходить под определение динозавров . М. К. Лангер и коллеги (2020) дали следующее определение, в которое включили первые описанные таксоны в каждой из основных подгрупп (кроме птиц): наименьшая клада, включающая игуанодона Iguanodon bernissartensis (птицетазовые), мегалозавра Megalosaurus bucklandii (тероподы) и цетиозавра Cetiosaurus oxoniensis (завроподоморфы) .
| Основной анализ |
Второй анализ, с использованием
матрицы данных из работы Langer et. al. , 2017 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
М. Барон и коллеги (2017) приводят альтернативное определение
|
|

Динозавров в целом можно описать как архозавров с . Многие группы доисторических животных, такие как ихтиозавры , мозазавры , плезиозавры , птерозавры и ранние синапсиды (в особенности диметродон ), неспециалистами часто ошибочно воспринимаются как динозавры, хотя научно не классифицируются как представители этой группы . Птерозавры состоят в дальнем родстве с динозаврами и входят вместе с ними в кладу орнитодир . Другие упомянутые группы, как и динозавры с птерозаврами, относятся к кладе завропсид , или рептилий, за исключением диметродона и других ранних синапсид, которые из современных животных наиболее близки к млекопитающим . Ни одно из этих животных не обладало выпрямленными задними конечностями, присущими настоящим динозаврам .
Динозавры были доминирующими наземными позвоночными мезозоя, особенно в юрский и меловой периоды. Остальные группы животных, за исключением некоторых других архозавров, в особенности наземных крокодиломорфов , в основном были сильно ограничены в своих размерах и занимаемых ими экологических нишах. Млекопитающие , к примеру, редко превышали размеры современной кошки, и нередко становились добычей плотоядных динозавров . Но имелись и исключения вроде крупного триконодонта Repenomamus giganticus , чей вес мог достигать отметки в 12-14 кг, причём такие габариты позволяли этому животному охотится и поедать детёнышей мелких динозавров, вроде детёнышей пситтакозавра . Среди гондванатериев существовал особо крупный представитель, найденный в маастрихтских отложениях Мадагаскара - Vintana sertichi , достигавшая веса в 9 кг и обладавшая хорошим обонянием, зрением и слухом. Динозавры представляют чрезвычайно разнообразную группу животных: по состоянию на 2018 год достоверно идентифицировано более 900 родов нептичьих динозавров и, по данным на 2016 год, 1124 вида . В 2016 году предполагаемое количество видов динозавров, существовавших в мезозойскую эру, было оценено в 1543—2468 . По оценке 2006 года, общее число родов, сохранившихся в палеонтологической летописи могло достигать около 1850, около 75 % из которых ещё не обнаружено . В ещё раннем исследовании, опубликованном в 1995 году, предполагается, что существовало около 3 400 родов нептичьих динозавров, из которых многие не сохранились в палеонтологической летописи . По данным Международного союза орнитологов , на январь 2021 года известно 10 806 современных видов птиц .
Некоторые из динозавров были растительноядными, в том числе употреблявшими в пищу семена, другие плотоядными и всеядными . Хотя предки этих животных являлись бипедальными животными, что было унаследовано многими их потомками, в том числе птицами, многие группы нептичьих динозавров были квадропедальными, а для продвинутых игуанодонтов и примитивных цератопсов, тиреофор и завроподоморф, был характерен или облигатный бипедализм, то есть они передвигались как на двух, так и на четырёх конечностях . Черепные модификации, например, рога и гребни, были присущи многим нептичьим динозаврам, а некоторые из них имели костяную броню. Хотя известно, что многие мезозойские динозавры были размером с человека или больше, современные птицы, как правило, обладают небольшим размером. На сегодняшний день считается, что динозавры населяли все континенты, а ископаемые остатки показывают, что они достигли глобального распространения по крайней мере в раннеюрскую эпоху . Современные птицы населяют большинство доступных сред обитания, от наземных до морских, и есть доказательства, что некоторые нептичьи динозавры (такие как микрораптор ) могли летать или, во всяком случае, скользить по воздуху , а другие, такие как некоторые спинозавриды , вели .

Хотя недавние открытия затруднили представление общепризнанного списка отличительных черт динозавров, почти все динозавры, описанные до сих пор, разделяют определённые модификации скелета, присущие также не относящимся к динозаврам архозаврам или более древним архозавроморфам , от которых те произошли. Хотя некоторые более поздние группы динозавров отличались несколько другими, дальнейшими модификациями этих признаков, они считаются типичными для динозавров; самые ранние динозавры обладали ими и передали своим потомкам. Такие модификации, происходящие от ближайшего общего предка некоторой таксономической группы, называются синапоморфиями данной группы .
Детальная оценка взаимоотношений архозавров подтвердила или обнаружила следующие двенадцать однозначных синапоморфий динозавров, некоторые из которых были известны ранее :
Несбитт представил ряд дальнейших потенциальных синапоморфий и отбросил ряд синапоморфий, предложенных ранее. Некоторые из них также присутствуют у силезаврид , которых Несбитт восстановил в качестве сестринской группы динозавров, в том числе большой передний трохантер, плюсневые кости II и IV почти равной длины, уменьшенная степень соединения между седалищной и лобковой костями, наличие кнемиального гребня на голени и восходящего на астрагале, а также многие другие . Тем не менее, в некоторых недавних филогенетических анализах силезавриды были восстановлены как града базальных птицетазовых динозавров .
Множество других скелетных особенностей характерны для всех динозавров. Однако, поскольку они присущи и другим группам архозавров , либо отсутствовали у некоторых ранних динозавров, эти особенности не считаются синапоморфиями. Например, как и большинство диапсид , динозавры имеют две пары (отверстия в черепе за глазницами ) и, как представители архозавров, обладают дополнительными отверстиями в передней части черепа и нижней челюсти . Кроме того, известно, что некоторые особенности, которые раньше считались синапоморфиями, возникли до появления динозавров или отсутствовали у самых ранних динозавров и независимо развились у различных групп динозавров. К ним относятся удлинённая лопатка, крестец, состоящий из трёх или более сросшихся позвонков (три встречаются у некоторых других архозавров, но у герреразавра их только два) и перфорированная вертлужная впадина или тазобедренный сустав с отверстием в её центре внутренней поверхности (к примеру, закрыто у сатурналии ) . Другая трудность определения чётко выраженных особенностей динозавров состоит в том, что ранние динозавры и другие архозавры позднего триаса часто плохо известны и во многих отношениях были похожи; эти животные иногда неправильно идентифицировались в литературе .

Задние конечности динозавров, как у большинства современных млекопитающих , выпрямлены, но отличаются от таких у большинства других рептилий, чьи конечности растягиваются по обе стороны . Это положение обусловлено развитием углубления в области таза, обращённого вбок (обычно открытого углубления и соответствующей обращённой внутрь головки на бедре ). Их вертикальное положение позволяло ранним динозаврам легко дышать во время движения, что, вероятно, способствовало выносливости и уровням активности, которые превышали таковые у «ползучих» пресмыкающихся . Вертикальные конечности, вероятно, также помогали поддержать развитие большого размера, уменьшая изгибающие напряжения на конечностях. У некоторых не относящихся к динозаврам архозавров, включая , также были прямые конечности, но это было достигнуто с помощью конфигурации тазобедренного сустава, в которой вместо выступа бедренной вставки на углублении на бедре, верхняя тазовая кость поворачивалась, формируя нависающий выступ .

Рептилии обитали на Земле ещё до появления динозавров. Лапы у них были расположены по бокам туловища, как у большинства современных ящериц. Около 300 млн лет назад во время каменноугольного периода произошло глобальное потепление , что, по мнению учёных, уничтожило тропические леса и вызвало эволюционный взрыв среди рептилий. Каждая популяция оказалась запертой в небольшой области обитания, и каждая начала развиваться по-своему, что и привело к увеличению разнообразия .
Затем появились предки динозавров — архозавры («господствующие ящеры»). Появление архозавров шло параллельно с массовым (но не полным) вымиранием терапсид на границе перми и триаса (см. Массовое пермское вымирание ). В начале триасового периода возникло множество новых видов.
Важнейшие продвинутые признаки ранних динозавров, которые отсутствовали у большинства текодонтов , были связаны с положением тела и характером локомоции . Задние конечности становятся практически вертикальными, а их длина по сравнению с более короткими передними конечностями позволяет делать выводы о хождении на двух ногах ( бипедальности ) .
Высшие формы терапсид по своей организации были очень близки к однопроходным млекопитающим и, по некоторым предположениям , имели молочные железы и шерсть . Внезапный упадок тероморфов и расцвет «настоящих ящеров» — одна из самых интригующих загадок палеонтологии .
В настоящее время невозможно определить точный момент, когда появились первые на Земле динозавры. Считается, что динозавры отклонились от своих предков архозавров спустя примерно 20 миллионов лет после массового пермского вымирания, которое уничтожило приблизительно 95 % всей жизни на Земле . Например, двуногий архозавр маразух из среднего триаса очень близок по строению к ранним динозаврам, в то же время у маразуха отсутствуют характерные особенности, присущие для всех динозавров .
На сегодняшний день остатки самых древних динозавров имеют возраст около 230 миллионов лет. Однако в 2010 году группа американских, немецких и южноафриканских палеонтологов представила в научном мире совместную работу, согласно которой первые динозавры населяли Землю на несколько миллионов лет раньше, чем это было принято считать. Свои выводы они сделали, проанализировав остатки новой динозаврообразной рептилии Asilisaurus kongwy , которая является родственным динозаврам видом, жившим 245—240 млн лет назад. Учёные говорят, что уже в это время различия между базальными динозавриформами и самыми ранними динозаврами стали существенными, следовательно, общий предок динозавров отделился от группы орнитодир значительно раньше, чем считалось прежде .
По одной из гипотез, оба отряда динозавров впервые появились на территории современной Южной Америки и затем распространились по всей Пангее в течение триасового периода с 230 до 199 миллионов лет назад .

Одним из самых ранних известных на сегодняшний день динозавров считается ставрикозавр , живший 228 млн лет назад, частичный скелет которого был обнаружен в отложениях среднего триасового периода на территории современной Бразилии . Большая берцовая кость длиннее бедренной , что характерно для бегающих животных, кости конечностей полые . В 1963 году был описан герреразавр , обнаруженный в отложениях позднего триаса на северо-западе Аргентины . Из-за фрагментарности остатков точная его классификация была не определена. Лишь после того, как в 1988 году был обнаружен более полный скелет с черепом, он в основном стал классифицироваться как примитивный теропод . Обнаруженный на территории современной Мексики вид Tawa hallae подтвердил связь герреразавра с более поздними тероподами (хищными динозаврами) . Через четыре года после открытия герреразавра, в Аргентине был найден скелет ещё одного древнего динозавра — эораптора , жившего 235—228 млн лет назад. Внешне он был похож на других рептилий, но особенности костей и зубов позволяют отнести его к примитивным тероподам . Палеонтологи полагают, что эораптор напоминает общего предка всех динозавров . Если эти предположения верны, то основываясь на его чертах, можно говорить, что первые динозавры были маленькими двуногими хищниками . Открытия примитивных подобных динозаврам орнитодиров , таких как маразух и , поддерживают эти предположения; анализ окаменелостей данных рептилий действительно говорит об их малых размерах и хождении на двух ногах.
Самым ранним из известных птицетазовых динозавров является пизанозавр ( Pisanosaurus ), обнаруженный в породах верхнего триаса на территории современной Аргентины. Лучше всего изучен род лесотозавр из юрских отложений Южной Африки. Как и все другие ранние птицетазовые динозавры, он достигал в длину почти одного метра. Строение задних конечностей характерно для бипедальных животных, большая берцовая кость значительно длиннее бедренной. Некоторые части скелета сходны с таковыми у ранних теропод, но тазовые кости и строение черепа и зубов уже имеют признаки птицетазовых динозавров .
На самых ранних этапах эволюции динозавров земная среда была заселена различными видами архозавров и терапсидов, такими как этозавры , цинодонты , дицинодонты , орнитозухиды , ринхозавры и другие. По всей видимости, в конце триаса динозавры выдержали долгую конкуренцию с данными животными, прежде чем победили и достигли расцвета. Некоторые учёные, однако, полагают, что господствующее положение динозавров было получено не столько превосходством над другими животными, сколько случайностью. В карнийском веке динозавры не показывают образец устойчивого увеличения в разнообразии и числе, как то должно было иметь место, если бы они, конкурируя, занимали ниши других групп животных. Вместо этого они были очень редки, составляя только 1—2 % от общей животной фауны. Лишь в норийском веке , после исчезновения нескольких других групп, они стали существенными компонентами фауны, представляя уже 50—90 % живых существ . Около 200 миллионов лет назад произошло так называемое триасовое вымирание . Это событие окончательно освободило экологические ниши, открыв возможность развиться, начиная с юрского периода , таким группам, как крокодиломорфы , птерозавры, млекопитающие, черепахи и динозавры .
Статистические исследования, основанные на имеющихся палеонтологических материалах, предполагают, что разнообразие динозавров, то есть число разновидностей, увеличилось в позднем меловом периоде .
Однако в июле 2008 года Грэм Т. Ллойд и его коллеги выдвинули утверждение, что это очевидное разнообразие является иллюзией, вызванной тем, что породы позднего мелового периода изучены в большей степени, чем породы юрского и триасового периодов. Вместо этого они написали статью о том, что в позднем меловом периоде существенной диверсификации подверглись только две группы динозавров — гадрозавры и цератопсы . В среднем меловом периоде цветущие покрытосеменные растения стали большей частью наземных экосистем, в то время как ранее доминировали голосеменные , такие как хвойные породы. Копролиты (окаменелый навоз) динозавров указывают на то, что в то время как некоторые ели покрытосеменные растения, большинство растительноядных динозавров питалось, главным образом, голосеменными растениями. В это же время растительноядные насекомые и млекопитающие развивались очень быстро и разносторонне, чтобы использовать в своих интересах новый тип пищи растительного происхождения. Ящерицы, змеи, крокодилы и птицы также разносторонне развивались в то же самое время. Ллойд и его коллеги предположили, что отказ нептичьих динозавров от разностороннего развития в свете изменения экосистемы обрекал их на исчезновение .
Со времени открытия первого динозавра их остатки были найдены на всех континентах планеты , включая Антарктиду (в июле 2022 года в журнале Science Advances международной группой ученых была опубликована статья, в которой приведены доказательства, что триасовые динозавры могли переживать морозы и даже обитали в полярных зонах ).
На сегодняшний день считается, что динозавры населяли все континенты, а ископаемые остатки показывают, что они достигли глобального распространения по крайней мере в раннеюрскую эпоху (современные птицы населяют большинство доступных сред обитания, от наземных до морских, и есть доказательства, что некоторые нептичьи динозавры (такие как микрораптор ) могли летать или, во всяком случае, скользить по воздуху, а другие, такие как некоторые спинозавриды , вели полуводный образ жизни).
На территории России случаи находок динозавров исчисляются единицами. Окаменелые кости находили лишь в Забайкалье ( амурозавр ) и на юге России . В Приамурье находится два довольно крупных кладбища динозавров: в Благовещенске и в Архаринском районе ( олоротитан ).
Во времена жизни динозавров их среда обитания ( окружающая среда ) достаточно сильно отличалась от современной — были иными и температура , и влажность и скорость вращения Земли и пр. Количество кислорода в атмосфере в 6 раз превышало нынешнее.



На сегодняшний день открыто большое видовое разнообразие динозавров. Одни из них передвигались на двух ногах, другие были четвероногие, а такие как Ammosaurus и игуанодон , могли легко передвигаться как на двух, так и на четырёх ногах. Многие из них обладали характерными особенностями, такими как костяная броня ( анкилозавр ) и роговые пластины ( стегозавр ) по всему туловищу, рога ( трицератопс ) или гребни на головах ( паразауролоф ).
Хотя динозавров часто представляют как животных больших размеров, многие из них были ростом с современного человека, а некоторые не превышали размеров современных кошек. В настоящее время одним из самых маленьких динозавров является Hesperonychus elizabethae , живший в Северной Америке и описанный в 2009 году . В длину он был 50 см, весил около 2 кг. Питался, по всей видимости, насекомыми, мелкими млекопитающими и детёнышами динозавров. В 2008 году был описан другой вид динозавра, Albertonykus borealis , также обитавший в Северной Америке и, возможно, достигавший меньшего размера, чем Hesperonychus , однако официально сообщать о каких-либо фактах палеонтологи не спешат, поскольку обнаруженных следов мини-динозавра недостаточно для серьёзных заявлений.


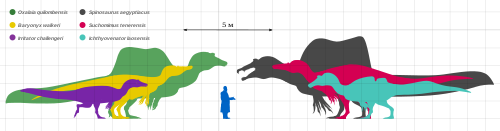
По современным оценкам, одним из самых крупных плотоядных животных, живших на Земле, является спинозавр . По расчётам на основе очень крупного фрагмента верхней челюсти, в длину он достигал около 15 метров. Вдоль спины у него была растянута кожа в виде паруса высотой около 1,8 метра.
Другими крупными тероподами являются гиганотозавр и кархародонтозавр , хорошо сохранившиеся образцы которых достигают в длину не менее 12 метров, а фрагментарные образцы могут указывать на животных крупнее 13 метров. Самый известный в современной культуре хищный динозавр — тираннозавр — также является одним из потенциально крупнейших тероподов, наибольший из его полных скелетов достигает длины 12,3 метров, а по некоторым данным, некоторые фрагментарные образцы и вовсе могли достигать длины более 14 метров .
В 1970 году в Монголии были раскопаны передние конечности дейнохейруса длиной почти 2,5 метра. Ввиду небольшого количества остатков, общая длина динозавра неизвестна. По найденным передним конечностям учёные предполагали, что в длину дейнохейрус мог достигать 20 метров. В настоящее время, однако, эти оценки были скорректированы до примерно 10 метров.
Четвероногие травоядные зауроподы и вовсе регулярно достигали длины 25—35 метров ( диплодоки , аргентинозавр , суперзавр , завропосейдон ). Высказаны предположения о существовании ещё более крупных зауропод , но их существование пока остаётся под сомнением.
Динозавры соединяли в себе свойства нескольких ныне обособленных групп животного царства: ящериц, крокодилов, млекопитающих и птиц. Их черепа в целом устроены по типу ящериц, но каждый зуб помещается в отдельной ячейке, как у крокодилов; с ними же сближает динозавров и кожистый или костяной панцирь, которым было покрыто туловище у некоторых родов. Со млекопитающими динозавров сближает строение трубчатых костей с широкими каналами, наполненными мозговым веществом, строение суставов пальцев и крестцовой кости (что, несомненно, является не более чем примером конвергентной эволюции в нишах крупных и активных наземных животных); наконец, по строению таза и задних конечностей они более всего приближаются к птицам.
Передвигались динозавры на вертикально расположенных конечностях, опираясь только на пальцы ( пальцехождение ), подобно большинству современных млекопитающих , чем отличались от современных рептилий , чьи конечности расположены по бокам туловища . Такое расположение конечностей произошло из-за развития скелетных особенностей в области таза в процессе эволюции . Их вертикальное положение позволило динозаврам при движении легко дышать и вероятнее всего повышало их уровень выносливости и активности. По этим параметрам они превзошли ползающих рептилий . Вертикальные конечности, вероятнее всего, также помогли поддержать эволюционное развитие до больших размеров, уменьшая нагрузку на сгибающиеся части конечностей . Некоторые «нединозавры» из других групп архозавров , таких как , также имели вертикальные конечности, но добились этого путём изменения строения тазобедренного сустава .

Ископаемые скелеты динозавров почти всегда являются единственным палеонтологическим материалом, по которому учёные пытаются воссоздать внешний облик древних ящеров. Характерной особенностью скелетов, разделяющей динозавров на две основные группы, является строение тазовых костей . У ящеротазовых динозавров подвздошные кости направлены вверх, где они соединяются с крестцом, седалищные — вниз и назад, а лобковые направлены вперёд и вниз. У птицетазовых динозавров седалищные и подвздошные кости устроены так же, а лобковые кости имеют две ветви, направленные в разные стороны: одна — вперёд, а вторая — назад, параллельно седалищным костям.
В настоящее время открытия большого видового разнообразия привели к тому, что трудно представить универсально согласованный список отличительных черт динозавров; почти всех обнаруженных на сегодняшний день динозавров разделяют определённые изменения по отношению к наследственному от архозавров скелету . Хотя некоторые более поздние группы динозавров показали признаки дальнейшего изменения общих черт, они считаются типичными динозаврами; такие общие черты между различными таксономическими группами называются синапоморфией .
Синапоморфия динозавров включает удлинённый гребень на плечевой кости или верхней кости руки для возможности крепления дельтопекторальных мышц; выступ в задней части подвздошной кости или главной тазовой кости ; голень или кость голени, отличающаяся широким нижним краем, а также возрастание проекции на таранной кости , формирующую нижнюю часть голеностопного сустава .
Существует множество других скелетных особенностей динозавров. Однако, поскольку они либо были общими для других групп архозавров или не присутствуют у всех ранних динозавров, эти признаки не считаются синапоморфией. Как и все архозавры , динозавры имели диапсидный тип черепа с двумя, а не с одной, как у других пресмыкающихся, височными впадинами позади глазниц . Кроме того, несколько характерных черт могли считаться синапоморфией, которые самостоятельно развивались у различных групп динозавров, но отсутствуют у ранних видов динозавров. Они включают удлинение лопатки; в тазовом поясе у динозавров был сложно устроенный крестец из трёх и более сросшихся позвонков (также три сращённых позвонка можно найти у некоторых других архозавров, в то время как у герреразавра их было два) и прободенная вертлужная впадина, куда входила головка бедренной кости. Эти анатомические особенности сделали динозавров самыми подвижными из сухопутных обитателей мезозоя .
Скелетные особенности, разделяющие динозавров на две группы, представляют собой ещё одно важное для палеонтологов доказательство явного родства между птицами и динозаврами. Области сходства скелетов птиц и динозавров включают в себя в основном строение шеи, конечностей и грудной пояс. Сравнение их скелетов путём кладистического анализа укрепляет аргументы в пользу их родства.


Восстановить мускулатуру и внутренние органы нептичьих динозавров учёным помогают знания о современных птицах и рептилиях. Также некоторые сведения можно почерпнуть, исследуя отпечатки динозавров. Образцы мягких тканей, таких как мышцы и внутренние органы, редко сохраняются в окаменелостях, и потому учёные спорят относительно анатомического строения динозавров.
В 1988 году в Италии был найден скелет мелкого хищного динозавра сципионикса , предполагаемый дыхательный аппарат которого по отпечаткам сходен с тем, который есть у нынешних крокодилов, и заметно отличается от птичьего. Некоторые признаки, такие как наличие « », присутствующего у крокодилов и отсутствующего у птиц, могли свидетельствовать о том, что сципионикс был эктотермным, то есть холоднокровным, хотя и способным поддерживать степень активности и потребления кислорода на более высоком уровне, чем у большинства современных пресмыкающихся . Впрочем, поскольку в настоящее время считается, что «печёночный поршень» достался современным крокодилам от теплокровных предков и изначально развивался именно как атрибут эндотермии, данная точка зрения может считаться малообоснованной . Более того, предполагаемые отпечатки мягких тканей сципионикса были сильно деформированы и другие авторы, которые изучали их после этого, сделали вывод что по этому образцу невозможно определить, сохраняет ли его печень правильную форму, и способна ли она действовать как часть печёночно-поршневого механизма крокодилов . Напротив, были предоставлены доказательства того, что некоторые, если не все динозавры обладали дыхательным аппаратом с воздушными мешками, подобно современным птицам .
В конце апреля 2000 года вызвала сенсацию находка якобы окаменелого сердца птицетазового динозавра тесцелозавра . Продвинутое строение сердца свидетельствовало о теплокровности ящера, однако «сердце» было окружено грубым песчаником красно-коричневого цвета со включением гравия, что свидетельствует о высокой скорости потока при заполнении грудной клетки осадочным материалом и о сопутствующей этому процессу окислительной среде с малыми шансами на литификацию мягкого органического вещества; кроме того, «сердце» находится снаружи грудной клетки и, вероятно, на самом деле является обычной конкрецией .
Также мало известно о нервной системе динозавров. На сегодняшний день было обнаружено несколько окаменевших мозговых полостей динозавров, в которых когда-то размещался мозг . По ним учёные предполагают, что мозг динозавров был похож на мозг современных рептилий и птиц.

Серия открытий, сделанных в 1990-х гг. в провинции Ляонин на севере Китая показала, что ряд мелких теропод обладали перьями. Первым найденным ископаемым динозавром со следами перьев на некоторых частях тела стал синозавроптерикс . Его «оперение» было образовано пуховыми перьями. Позднее в 1997 году был открыт каудиптерикс , внешним видом напоминавший птицу. Судя по окаменелостям, большая часть его тела была покрыта мелким пухом, а на хвосте и передних конечностях росли длинные жёсткие перья .
Обладал перьями и микрораптор , который подобно археоптериксу , являет собой важное доказательство родства динозавров и птиц. Микрораптор имел длинные с асимметричными бородками, которые образовывали крылоподобные поверхности не только на передних конечностях и хвосте, но, что удивительно, и на задних конечностях. Это подтолкнуло исследователей к тому, чтобы предположить, что он мог планировать, используя все четыре конечности . В южной части пустыни Гоби был открыт небольшой оперённый динозавр — мононик , каждая кисть которого имела только один крупный коготь.

На сегодняшний день самыми ранними из известных науке оперённых динозавров являются тероподы педопенна (около 164—159 млн лет назад) и анхиорнис (160—155 млн лет назад) и представитель ящеротазавых Tianyulong (ок. 158 млн л. н.). К этому же времени относится и другой динозавр, Epidexipteryx , с характерными лентообразными перьями на хвосте, напоминающими некое подобие павлиньих. По мнению учёных это свидетельствует о том, что в этот период привычное оперение ещё не успело сформироваться, в результате чего и появляются динозавры с необычным внешним видом .
В сентябре 2010 года группа учёных из Национального университета Мадрида описали новый вид динозавра — конкавенатор , на предплечевой кости которого были обнаружены характерные для перьев бугорки, интерпретируемые как точки прикрепления перьев. В результате этого учёные выдвинули предположение, что некие подобия перьев начали появляться уже у ранних представителей клады Neotetanurae , живших 171—165 миллионов лет назад, и эти данные отодвигают в прошлое момент появления перьеподобных структур .
Также среди учёных есть мнение, что некое подобие перьев уже имели самые первые динозавры. Эти предположения были выдвинуты в связи с обнаружением волосовидного пуха у Tianyulong и пситтакозавра , которые относятся совсем к другой группе динозавров — птицетазовым — в отличие от Neotetanurae , принадлежащих к ящеротазовым. Обе эти ветви динозавров разделились на раннем этапе своего развития. Сторонники этой гипотезы не исключают такой возможности уже потому, что перья могли просто не сохраниться вместе с ископаемыми остатками динозавров .
Палеонтологи обнаружили в Китае прекрасно сохранившиеся остатки пернатого динозавра размером с серую ворону ( Caihong juji ), часть перьев которого переливалась, как у попугаев, а другие были тёмными. Описание находки опубликовано в журнале Nature Communications.
В 1993 году был обнаружен почти полный скелет тесцелозавра ( Thescelosaurus ), в грудной клетке которого была обнаружена трёхмерная структура, в результате чего была выдвинута гипотеза, что это ископаемое сердце динозавра . В 1998 году в Италии был найден скелет сципионикса ( Scipionyx ). Большинство костей его скелета сохранилось в идеальном состоянии, также сохранились отпечатки мышц, кишечника, печени и трахеи .
В 2005 году в журнале Science была опубликована статья палеонтолога Мэри Швайцер с соавторами, в которой сообщалось о выделении коллагена из костей тираннозавра , жившего 68 млн лет назад. Обнаружилось большое количество железосодержащих органических молекул, отсутствующих в тканях современных животных. Поэтому сохранность коллагена авторы объяснили его связыванием с ионами железа , что приводит к устойчивости к бактериальному и ферментному разложению. Также остался незамещённым в костях фосфат кальция (биогенный апатит). Предполагается, что это вещество сохранилось до наших дней благодаря присутствию в окружающей породе кальцита. В нейтральных условиях в присутствии кальцита апатит не растворяется . Эта публикация вызвала серьёзную критику и в 2008 году вышла публикация Томаса Кайе с соавторами. В ней было показано, что обнаруженные Швайцер артефакты могут оказаться современными биоплёнками . Однако в следующем году вышла ещё одна статья Швайцер с соавторами, в которой сообщалось об обнаружении коллагена и мягких тканей в окаменелостях другого динозавра — брахилофозавра , жившего 80 млн лет назад. После деминерализации костей под микроскопом были отчётливо видны кровеносные сосуды (иногда с округлыми красноватыми включениями, напоминающими эритроциты ), а также остеоциты . Выделенные структуры очень похожи на аналогичные структуры современного страуса после аналогичной обработки, имитирующей деминерализацию .
Из костей брахилофозавра было выделено восемь пептидных последовательностей общей длиной 149 аминокислот . Филогенетический анализ с использованием аминокислотных последовательностей коллагена тираннозавра и брахилофозавра подтвердил близкое родство птиц с динозаврами .
Вопрос о том, были ли динозавры теплокровными или холоднокровными животными, является дискуссионным, однако в последнее время учёные всё больше склоняются к теплокровности. Сейчас зачастую дискуссии палеонтологов и палеобиологов касаются не самого наличия у динозавров признаков теплокровности, а вопроса, какие динозавры обладали постоянной температурой тела, а какие — нет.
Так, некоторые оценки интенсивности метаболизма динозавров показывают, что двуногие динозавры массой 20 кг и более в любом случае были теплокровными . Однако, спорным вопросом является способ поддержания температуры тела. Свидетельство того, что крупные динозавры были инерциальными эндотермами , получили в 2006 году Джеймс Гиллули, Эндрю Аллен и Эрик Чарнов. Исследовав рост костей крупных динозавров, они выяснили, что в начале жизни температура тела крупных динозавров составляла +25 °C , а по мере достижения массы в несколько сотен килограммов она значительно вырастала и достигала у самых крупных экземпляров +35 °C. В то же время у мелких динозавров с ростом температура тела не увеличивалась. Был сделан вывод, что по мере увеличения размеров возникает способность удерживать тепло.
О теплокровности некоторых видов говорят также остатки, найденные в Австралии , приполярном холодном районе на момент жизни особей.
В июле 2022 года в журнале Science Advances международной группой ученых была опубликована статья, в которой приведены доказательства, что триасовые динозавры могли переживать морозы и даже обитали в полярных зонах. Палеонтологи изучили следы динозавров на ледовых скалах Джунгарской впадины в Китае и сделали вывод, что ко времени триасово-юрского вымирания некоторые динозавры успели приспособиться к низким температурам, что позволило им выжить и адаптироваться к жизни в холодных районах Земли .
В 2010 году учёные из Музея естественной истории имени Филда ( Чикаго ) пришли к выводу, что появление клюва у динозавров было эволюционно значимо. Благодаря развитию клюва тероподы получили доступ к новым для себя продуктам питания. Этот вывод они сделали, обобщив данные о рационе тероподов. Хищники всегда распространены меньше, чем растительноядные. Поэтому, по мнению учёных, многим тероподам (например Caudipteryx zoui , Beipiaosaurus inexpectus ) и их родственникам приходилось довольствоваться вегетарианской диетой. Об этом свидетельствуют ископаемый навоз, содержимое желудка, зубы и прочее. Также исследователи выявили и другие анатомические особенности, которые связаны с растительной диетой, например, выпадение зубов и удлинение шеи .


Многие современные птицы социальны и живут стаями. Существует общепринятое мнение, что некоторые поведенческие особенности, которые распространены у птиц, а также у крокодилов (ближайших современных родственников птиц), были характерны и для вымерших групп динозавров. Выводы о поведении ископаемых видов динозавров сделаны, как правило, на основании поз тел ископаемых, обстановки окружающей среды, сравнения с поведением современных животных, занимающих сходные экологические ниши , а также при помощи компьютерного моделирования их биомеханики .

Некоторые динозавры обладали социальным поведением . На данный момент наиболее ранним из подобных динозавров являются майазавры ( Maiasaura , название дословно переводится как «ящеры-няньки») и пситтакозавры , жившие 130—100 млн лет назад . Находки позволяют сделать вывод, что уже у динозавров мелового периода существовал родительский инстинкт .
Динозавры вели как одиночный, так и групповой образ жизни. Параллельно идущие цепочки следов множества динозавров , а также совместные захоронения десятков особей одного вида, близких по возрасту (по-видимому, утонувших в болоте) показывают способность некоторых динозавров объединяться в крупные стада.
Также имеются доказательства того, что некоторые виды динозавров (например небольшой Oryctodromeus cubicularis из группы гипсилофодонтов ) выкапывали себе норы. Данная находка была сделана в 2005 году в штате Монтана . Палеонтологи обнаружили не только их норы, но и сами кости динозавров. Изучение костей показало, что динозавры по какой-то причине умерли в своём убежище, а потом оно оказалось засыпано, возможно, во время наводнения. В 2009 году в Австралии повторно были обнаружены норы динозавров. До открытия нор палеонтологи предполагали, что динозавры спали где-то под корнями деревьев и в других естественных убежищах. Теперь же учёные предполагают, что некоторые виды динозавров выкапывали себе норы как раз для спячки .

В 2011 году палеонтологи из Калифорнийского университета в Дэвисе установили, что многие динозавры охотились ночью. До этого среди учёных было распространено мнение, что динозавры были активны преимущественно днём. Палеонтологи благодаря специально созданной программе, исследовали 33 вида древних ящеров. На основе этих исследований, палеонтологи установили, что многие из динозавров вели ночной образ жизни. Среди них были выделены велоцирапторы , а также некоторые виды птерозавров .
Динозавры откладывали яйца, которые имели очень прочную скорлупу и по большому счёту ничем не отличались от яиц птиц и других рептилий. Большинство динозавров строили гнёзда для высиживания потомства.


Впервые ископаемые были найдены в 1859 году во Франции . Они принадлежали гипселозавру .
В целом, форма и размер яиц динозавров варьировали, обычно они имели округлую или удлинённую форму. Длина самого крупного яйца, найденного в Восточном Китае , составляла 45 см, и принадлежало оно, вероятно, теризинозавру . Крупнейшее захоронение ископаемых яиц было обнаружено в 1995 году также в Китае.
Гнёзда динозавров были обнаружены ранее — в 1923 году в пустыне Гоби . Они представляют собой неглубокие ямки в земле или невысокие возвышения округлой формы с углублением посередине. Откладывая яйца, самка, вероятно, приседала над гнездом. Яйца в таких гнёздах обычно располагались рядами или полукругом .
В 2010 году группа учёных из США и Аргентины представили доказательства того, что некоторые динозавры устраивали паровой обогрев кладок яиц, откладывая их рядом с геотермальными источниками .
В ряде случаев по ископаемым остаткам, содержащим пигмент, удаётся установить цвета окраски динозавров .

На протяжении большей части мелового периода Австралия и Новая Зеландия были соединены с Антарктидой, образуя полярный континент. Хотя климат в те времена был гораздо теплее, чем сегодня, обитавшие здесь динозавры были вынуждены приспосабливаться к суровым погодным условиям. Летом данную область круглые сутки освещало солнце, а на протяжении пяти месяцев в году царила полярная ночь . Большинство находок подобных динозавров было сделано на побережье штата Виктория (Южная Австралия) в так называемой Бухте динозавров. Все динозавры, найденные тут, жили в ранний меловой период . Большинство находок принадлежали небольшим травоядным орнитоподам — леэллинозаврам ( Leaellynasaurus ), квантасзаврам ( Qantassaurus ), а также к гадрозаврам . В данной области также были найдены остатки хищников — вида, принадлежащего к орнитомимозаврам , и вида из группы аллозавров . Возможно, что хищники и травоядные динозавры находились в данных областях летом, а зимой перекочёвывали в более тёплые края на север .
Малочисленность находок динозавров в Антарктиде частично вызвана тем, что около 98 % её поверхности сейчас находится подо льдом. Первая находка динозавров на территории Антарктиды была сделана в 1986 году . Было обнаружено несколько видов. Большинство из найденных остатков являются фрагментарными, из-за чего ряд из них до сих пор не получил научных названий. На острове Росса у северо-западной части Антарктиды, обнаружены остатки анкилозавров и динозавра из группы гипсилофодонтид . На найдены остатки динозавра из группы гадрозавров . В 1991 году в Антарктиде на склоне горы Керкпатрик нашли остатки прозавропода , а также теропода криолофозавра , достигавшего семи метров в длину и обладавшего гребнем на голове шириной 20 см .

Все нептичьи динозавры вымерли в конце мелового периода , около 66 миллионов лет назад. Было ли это вымирание постепенным или внезапным , в настоящее время — предмет споров; единой точки зрения по этому поводу нет.
Вымирание нептичьих динозавров явилось лишь частью так называемого «великого вымирания», имевшего место в то же время: вместе с динозаврами вымерли морские рептилии ( мозазавры и плезиозавры ) и летающие ящеры , многие моллюски, в том числе аммониты , белемниты и множество мелких водорослей. Всего погибло 16 % семейств морских животных (47 % родов морских животных) и 18 % семейств сухопутных позвоночных.
Согласно одной из гипотез, причиной «великого вымирания» стал удар астероида или кометы в районе мексиканского полуострова Юкатан (кратер Чикшулуб ) , в результате чего в течение 3—16 лет после падения астероида температура на планете могла снизиться на 26 °C . Существуют и другие гипотезы причин вымирания нептичьих динозавров .

Гигантские кости, находимые иногда в земле, в античности считали останками героев эпохи Троянской войны , в Средние века и вплоть до XIX в. — останками исполинов , о которых упоминается в Библии и которые погибли во время всемирного потопа ; на Дальнем Востоке их считали костями драконов и приписывали им целебные свойства.
В 1824 году президент Королевского геологического общества Уильям Баклэнд выступил с докладом о находке, сделанной в 1815 году в юрских сланцах Стоунзфилда (графство Оксфордшир ) и состоящей из нескольких костей и фрагмента «допотопного» животного. Прибегнув к помощи видного специалиста по сравнительной анатомии Жоржа Кювье , Баклэнд классифицировал находку как ископаемые остатки гигантской хищной ящерицы ( лат. sauria ) и, соответственно, назвал её мегалозавром — «огромным ящером».
В 1826 году Гидеон Мантелл , хирург из Льюиса (графство Суссекс ), действительный член Линнеевского общества , аналогичным образом представил в Геологическом обществе найденные им зубы ранее неизвестного вида, которому он дал название игуанодон (букв. «игуанозубый») за сходство зуба с зубом ящерицы игуаны . Он же в 1833 году описал гилеозаврa — представителя панцирных ящеров анкилозавров .



В 1842 году английский биолог Ричард Оуэн , констатировав несомненное сходство между этими тремя видами и их отличие от современных рептилий, выделил их в особый подотряд , назвав его Dinosauria («ужасные ящеры») .
Открытие в 1858 году в США хорошо сохранившегося скелета гадрозавра опрокинуло представление о динозаврах как о четвероногих животных, показав, что динозавры могли ходить на двух ногах. В последующие несколько десятилетий были открыты представители большинства основных групп динозавров; важная заслуга в этом принадлежит американским палеонтологам Гофониилу Маршу и Эдварду Копу , которые открыли и описали в общей сложности 142 новых вида, включая апатозавра и бронтозавра (впоследствии их отнесли к одному роду), диплодока и стегозавра , моноклона , трицератопса и др. Накопление материала привело к разделению динозавров на семейства птицетазовых и ящеротазовых ( 1887 ).
В рамках первой половины двадцатого века бо́льшая часть научного сообщества ошибочно полагала, что динозавры были громоздкими, вялыми животными. Большинство исследований, проводимых с 1970 года , однако, указали, что динозавры были активными животными с повышенным метаболизмом и многочисленными особенностями для социального взаимодействия.
В 1964 году находка дейнониха произвела новую научную революцию, так как по строению динозавра было ясно, что он передвигался относительно быстро, из чего следовал вывод, что он был теплокровным. Идея теплокровности заставляла пересмотреть старые представления не только о физиологии, но и о поведении динозавров, чему появились подтверждения в 1979 году , когда были получены доказательства родительского инстинкта и социального поведения ящеров (высиживание, защита и выкармливание детёнышей). Наконец, сравнение верхних конечностей дейнониха с крылом птицы заставило предполагать их близость и происхождение птиц от динозавров (или даже вообще принадлежность к этому надотряду), чему доказательством стало впоследствии открытие следов оперения у ряда динозавров. В 2005 году учёным удалось выделить коллаген из сохранившихся мягких тканей тираннозавра и использовать его химический состав как ещё одно доказательство родства динозавров с современными птицами .
Ряд учёных утверждает, что около трети описанных видов динозавров не существовали. За ранее неизвестных ящеров учёные принимали уже описанных динозавров на разных стадиях развития. Проведённое масштабное исследование другими учёными показало, что почти 50 % всех видов динозавров были названы неправильно .
В настоящее время наблюдаются две противоположно направленные тенденции в систематике динозавров. В то время как одни палеонтологи продолжают дробление существующих таксонов, выделяя по тем или иным признакам новые рода и виды из уже существующих, другие их коллеги ставят под сомнение корректность ранее описанных видов. Так в 2007 году Джек Хорнер ( англ. Jack Horner ) опубликовал статью, в которой высказывал, что Dracorex hogwartsia , Stygimoloch spinifer и Pachycephalosaurus wyomingensis являются тремя разными возрастными стадиями одного вида. В 2009 году Джек Хорнер при изучении черепа Nanotyrannus заключил, что этот динозавр на самом деле является молодым Tyrannosaurus , тем самым подтвердив интерпретацию черепа CMNH 7541 как ювенильного T. rex , предложенную А. К. Рождественским в 1965 году . В 2010 году в издании Journal of Vertebrate Paleontology вышла статья палеонтологов из Государственного университета Монтаны ( Montana State University ), где также говорится, что Triceratops и Torosaurus — различные стадии роста особей одного вида. В том же году другие учёные из Йельского университета описали новый род динозавров — Mojoceratops , кости которого раньше причисляли к Chasmosaurus .
Долгое время на территории России не удавалось найти скелетов динозавров. Ещё Отниел Чарльз Марш приехавший в Российскую империю в 1891 году с удивлением узнал, что в стране не найдено ни одной кости динозавра. Это было обусловлено в первую очередь тем, что в юрском и меловом периодах половину нынешней территории России покрывали мелководные моря. Ещё одна причина — отсутствие бедлендов , огромных пустынных районов, изрезанных ущельями и каньонами. Самой ранней находкой считается фрагмент стопы ящера, найденной в Читинской области палеонтологом Анатолием Рябининым в 1915 году . Он описал её под именем Allosaurus sibiricus . Позднее и в других местах стали находить остатки динозавров. На берегах Амура было найдено множество костей, была организована экспедиция, которая прямо перед революцией доставила в Санкт-Петербург более тонны ископаемых остатков. Из них собрали большой скелет, описав его как новый вид утконосого динозавра — . Однако позже стало ясно, что кости принадлежат разным видам и родам ящеров. В 1930-х годах японские палеонтологи выкопали в угольных копях Сахалина новые остатки ящера — Nipponosaurus sachalinensis .
Только в 1953 году палеонтологам по-настоящему повезло. На реке Кия близ села Шестаково геологам попался череп и неполный скелет небольшого, размером с собаку, пситтакозавра , которого назвали сибирским ( Psittacosaurus sibiricus ). Спустя полвека в Шестакове удалось найти ещё несколько костей позвонков завропода .
Начиная с 90-х годов изучение российских динозавров сильно продвинулось. Найден десяток крупных захоронений, удалось отыскать ценные остатки в ранее известных местах находок. В конце 1990-х годов в сопках возле Кундура при прокладке дороги геологи нашли полный скелет ящера, названного олоротитаном ( Olorotitan arharensis ). Данный динозавр примечателен тем, что он является одним из последних, обитавших на планете. Возле города Шарыпово в Красноярском крае были найдены скелет стегозавра и хищный динозавр — Kileskus aristotocus . На берегу Волги , недалеко от посёлка Сланцевый Рудник в Ульяновской области , было обнаружено семь позвонков, которые учёные отнесли к новому таксону титанозавров ( Volgatitan simbirskiensis ) .
Главные захоронения российских динозавров расположены за Уралом — в Кундуре , Благовещенске , Шестакове . Находки на берегу реки на Корякском нагорье считаются самыми северными обнаружениями динозавров на планете. Здесь найдены кости семи семейств и скорлупа яиц как минимум двух видов динозавров. Остатки меловых динозавров также обнаружены в Бурятии (местонахождения Муртой и Красный Яр) и Красноярском крае ( Большой Кемчуг ). Динозавры юрского периода найдены в Якутии (ручей ) и в Республике Тыва . В западной части России нет захоронений с целыми скелетами и черепами динозавров. Здесь, прежде всего в Поволжье и Белгородской области , попадаются в основном разрозненные остатки. В ста километрах от Москвы , у железнодорожной станции Пески найдены остатки хищных целурозавров (Coelurosauria) .

Сравнительная давность открытия и исследования динозавров , а также более чем полуторастолетняя история термина, привели к тому, что динозавром непосвящённые люди склонны называть любую доисторическую рептилию. Это неверно, так как история эволюции знает и другие большие группы ископаемых рептилий, доминировавшие на Земле за миллионы лет до появления динозавров — например, анапсиды и синапсиды триасового и пермского периодов (они исчезли в результате так называемого массового пермского вымирания , «открыв» тем самым дорогу для предков динозавров — архозавров ).
Наличие большого числа хорошо сохранившихся ископаемых остатков, включённых в экспозиции крупнейших палеонтологических музеев мира, также весьма поспособствовало знаменитости этой группы доисторических ящеров.
Таким образом, сравнительная изученность как самих динозавров, так и драматической истории их вымирания в конце мелового периода , а также впечатляющие размеры, обусловили распространённость их образа в массовом сознании, сделав динозавров популярными «героями» многочисленных произведений.
С точки зрения геральдической науки изображение нептичьего динозавра является естественным негеральдическим символом. Оно используется в символике тех местностей, где были найдены остатки динозавров.

О птицах в геральдике читайте в статье Птицы в геральдике .
{{
cite news
}}
:
Проверьте значение
|url=
(
справка
)
|
|
Некоторые
внешние ссылки
в этой статье
ведут на сайты, занесённые в
спам-лист
|