Interested Article - Дикинсонии

- 2021-12-18
- 1
Дикинсо́нии , или диккинсо́нии ( лат. Dickinsonia ) — один из наиболее характерных родов ископаемых животных эдиакарской (вендской) биоты . Обычно дикинсонии представляют собой двусторонне-симметричное рифлёное овальное тело. Современные учёные относят дикинсоний к животным , к группе проартикулят , однако существуют мнения, что они являются грибами или относятся к особому, ныне не существующему царству живой природы.
Видовое разнообразие

Всего описаны 9 видов дикинсонии:
- Dickinsonia costata Sprigg, 1947 typus
- Dickinsonia minima Sprigg, 1949
- Dickinsonia spriggi Harrington et Moore, 1955
- Dickinsonia elongata Glaessner et Wade, 1966
- Dickinsonia tenuis Glaessner et Wade, 1966
- Dickinsonia lissa Wade, 1972
- Dickinsonia brachina Wade, 1972
- Dickinsonia menneri (Keller, 1976) , повторно описаны Иванцовым в 2007 году (= Vendomia menneri Keller 1976 )
- Dickinsonia rex Jenkins, 1992
Из них действительными признаны лишь три :
- D. costata . Младшие синонимы: D. minima , D. spriggi и D. elongata . Отличается от других видов округлой формой тела.
- D. tenuis . Младшие синонимы: D. brachina , D. lissa и D. rex . Отличается от D. costata удлиненной овальной формой тела, более такими и многочисленными изомерами.
- D. menneri — небольшой организм около 8 мм в длину, напоминающий незрелую особь D. costata небольшим числом изомеров и хорошо очерченной головой, отличается от D. costata удлинённой формой.
Морфология
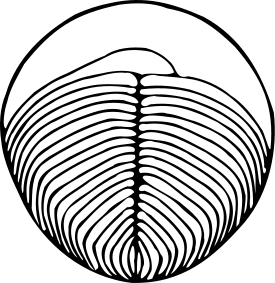
Особи от 1 мм до 1,4 м в длину овальной или змеевидной формы. Состоят из большого количества ленточных сегментов, выходящих из центрального гребня или желобка. Полусегменты (изомеры) обеих сторон сдвинуты относительно друг друга на половину ширины, образуя симметрию скользящего отражения .
Самые крупные экземпляры были обнаружены в эдиакарских отложениях формации Вонока в Южной Австралии. На Зимнем берегу Белого моря в Архангельской области были найдены окаменелые экземпляры дикинсонии длиной до 55 см .
Сегменты дикинсонии описываются как «пневматики», заполненные жидкостью под давлением выше, чем давление окружающей среды . На некоторых экземплярах замечены следы, интерпретируемые как продольные мышечные волокна и медиальная кишка, но эта интерпретация не общепринята.
Полагают, что внутренности дикинсонии содержат веретенообразные волокна толщиной 0,5—1 мм .
Находки окаменелостей
Таксон Dickinsonia впервые описан , первооткрываетелем эдиакарской биоты в Австралии . Назван им в честь , директора горнорудной промышленности Южной Австралии и главы правительственного департамента, где работал Спригг.
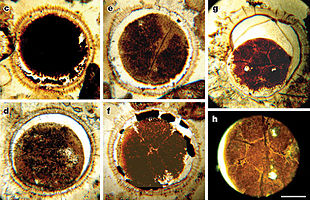
Отпечатки дикинсоний обнаруживают в кварцевых мелкозернистых песчаниках и алевролитах позднего эдиакарского периода . Наиболее известны находки из Эдиакары и других местностей хребта Флиндерс в Южной Австралии . Дикинсонии обнаружены в вендских отложениях на Восточно-Европейской платформе . На побережье Белого моря ( Зимний берег ) обнаружили дикинсоний отличной сохранности (беломорский стиль сохранности). У некоторых беломорских образцов сохранилась тонкая органическая плёнка. Дикинсонии обнаружены на Среднем Урале в России (сылвицкая серия), в Подолье на Украине , в Мадхья-Прадеш в Индии (группа Бхандер) и из ущелья Янзцзы в Южном Китае ( ) . Возраст окаменелостей оценивается в 560—555 млн лет .
Находки дикинсоний являются «устойчивой» окаменелостью, в отличие от других эдиакарских организмов они сохраняются как вогнутый отпечаток на подошве вышележащего слоя (негативный гипорельеф). Если сохранился и верхний, и нижний отпечаток, они обычно отстоят по вертикали на расстояние около 3 мм с наиболее сильным рифлением на верхнем отпечатке, что позволяет предположить, что рельеф имелся только на спинной поверхности организма.
В окаменелостях дикинсоний, найденных в Лямце Ильёй Бобровским и др. из Национального университета Австралии в Канберре , и в окаменелостях андивы ( Andiva ) с Зимнего берега обнаружены органические вещества (насыщенные производные стерана и моноароматические стероиды ), в которые превращаются в процессе фоссилизации эукариотические стеролы — компоненты клеточных мембран эукариот. В ископаемых дикинсониях были найдены в основном стеролы с углеродным скелетом, содержащим 27 атомов углерода ( , C 27 ), характерные для организмов многоклеточных животных, тогда как грибы и лишайники содержат C 28 ( ), зелёные водоросли — C 29 ( ), а крупные протисты — смесь C 27 , C 28 , C 29 (некоторые также C 30 ); этот анализ подтвердил гипотезу о принадлежности дикинсонии к царству животных .
Следы
Найдены дугообразные следы дикинсонии, отнесённые к ихнотаксону , однако их интерпретация не бесспорна. Это могли быть слизистые выделения организма на подстилающие микробные маты , возможно, с целью переваривания . Некоторые исследователи интерпретируют эти окаменелости как следы перекатывания по дну какого-то предмета или организма под действием потока воды либо отпечатки, оставленные группой лишайников или грибов (наподобие « ведьминых кругов ») . Однако в некоторых случаях следы перекрывают друг друга. Наблюдаются дорожки, сформированные, вероятно, кишечными выделениями, подтверждающие версию, что следы оставлены в процессе питания. К тому же нарушения структуры осадочных пород, вызванные падением или перекатыванием по дну, не обнаружены .
Взаимодействие окаменелостей
Не обнаружено ни одного случая, когда бы окаменелости отдельных организмов перекрывались. Вокруг каждого организма наблюдается своеобразная «полоса отчуждения». Организмы, находящиеся в непосредственной близости друг от друга, часто деформированы, чтобы не нарушить её .
Внутренняя анатомия

Некоторые окаменелости дикинсоний сохранили элементы внутренней анатомии. Предполагается, что они представляли собой тракт, обеспечивавший переваривание пищи и распределение питательных веществ по организму . На одном из месторождений Зимнего берега в Архангельской области найдено более двадцати отпечатков дикинсоний со следами желудочно-кишечного тракта .
Тафономия
Дикинсонии, как и другие обитатели эдиакарской биоты , не имели минерализованого скелета (раковины или панциря), их тело состояло только из мягких тканей, подобно современным медузам или слизням. По этой причине вендские окаменелости представлены только отпечатками на мелководных песчано-глинистых отложениях. Возникновение отпечатков связывают с лавинообразным накоплением осадков во время шторма или подводного оползня. Когда участок дна накрывался новым осадочным слоем, на подошве последнего образовывались отпечатки погребённых им живых организмов. Различают следующие типы отпечатков по рельефу :
- негативные, вдавленные в слой — представляют собой отпечатки верхней части животных, накрытых и сдавленных осадочным слоем;
- позитивные, возвышающиеся над слоем — представляют собой вмятины на грунте, заполненные вышележащими осадочными породами. Обычно это следы животных или отпечатки нижней части их тел.
Благодаря тому, что разные части тела имели разную плотность и разлагались с разной скоростью, в рельефе отпечатков обнаруживается не только топография наружной поверхности организма, но и элементы его внутреннего строения. Более стойкие ткани и органы образовывали выступы, менее стойкие — впадины .
Многочисленные отпечатки мягкотелых животных являются отличительной особенностью ориктоценозов эдиакарских отложений. В эдиакарской биоте отсутствовали падальщики, измельчавшие мёртвые тела, и биотурбаторы (роющие животные), разрушавшие отпечатки на мягком грунте. Мёртвая органика разлагалась только благодаря жизнедеятельности микробов .
В тех образцах, где дикинсония сложена или согнута, она демонстрирует отсутствие разрушений, характерных для хрупких структур .
Толщина найденных образцов мало зависит от линейных размеров, что характерно скорее для лишайников , лиственных растений или грибов , чем для мягкотелых животных . Полагают, что дикинсонии были более жёсткими, чем черви или медузы благодаря гидростатическому скелету .
В обнаруженных скоплениях организмов присутствуют особи различных размеров, что свидетельствует об их захоронении в естественных условиях. В скоплениях, образованных наносами водяных потоков преобладали бы особи меньших размеров . Обнаружение некоторых экземпляров на вершинах осадочных структур говорит об их надёжном прикреплении к дну в момент захоронения .
Дикинсоний обнаруживают в осадочных слоях толщиной от 8 мм. Это свидетельствует о том, что их толщина с учётом сдавливания не превышала 1 см.
Экология
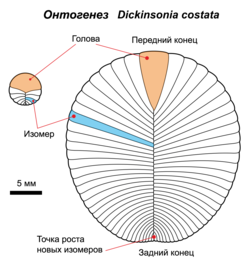
У дикинсоний наблюдается неограниченный изометрический рост — увеличение геометрических размеров с сохранением пропорций до тех пор, пока организм не будет захоронен осадочными породами или убит каким-либо другим способом . Большую часть жизни они, вероятно, проводили в неподвижности, прикреплёнными к донным осадкам, однако могли передвигаться с места на место. Тип прикрепления к субстрату, возможно, схож со сращением устриц, ризоидами лишайников или подобием подземной грибницы . Сохранность организмов свидетельствует, что несущими элементами окаменелостей были скорее биополимеры типа кератина , чем хрупкие минералы типа кальцита или посмертная пиритизация тела.
Родовое сходство
Родственные отношения дикинсонии с современными типами животных неясны. Различные исследователи относят её к медузам , кораллам , морским анемонам , , многощетинковым или ресничным червям , грибам , лишайникам и даже близким предшественникам хордовых .
Реталлэк сделал попытку проследить родственные отношения дикинсонии, используя сходство процесса разложения останков и предположил, что организмы эдиакарской биоты были родственны лишайникам , однако его взгляды подверглись резкой критике . Тем не менее, режим разложения эдиакарских организмов действительно более сходен с процессом разложения лиственных растений, грибов и лишайников, чем мягкотелых животных, которые в процессе распада сжимаются и деформируются .
Долгие споры о родственных связях эдиакарских организмов привели к взгляду (который был впервые высказан Раннегаром в 1958 году ), что они относятся к особому царству живой природы, «вендобионты» или «вендозоа», которые вымерли на границе кембрийского периода и не оставили потомков среди современных типов организмов . Другие вендобионты ( ёргия , Marywadea ) имеют много сходных черт с дикинсонией и, видимо, состоят с ней в родстве. Основные отличия дикинсонии от животных современных типов следующие :
- изометрический рост, то есть рост с сохранением пропорций организма, тогда как у животных современных типов пропорции тела различаются у детёнышей и взрослых особей ;
- своеобразный тип симметрии, сходный с билатеральной симметрией современных организмов, однако со сдвигом правых и левых полусегментов относительно друг друга. М. А. Федонкин обозначил этот тип симметрии заимствованным из математики термином « симметрия скользящего отражения ». У современных животных скользящая симметрия встречается крайне редко (один из примеров — расположение мышечных элементов — миомеров и миосепт — у ланцетника ) .
Отмечено, что симметрия скользящего отражения характерна для листоподобных органов папоротника, в связи с чем некоторые исследователи относят дикинсонию и похожие на неё организмы (например, чарнию ) в таксон .
Общепризнано, что дикинсонии могли передвигаться, о чём свидетельствуют их следы на бактериальных матах , оставленные в процессе питания . Однако нет бесспорных свидетельств наличия у дикинсоний рта , кишечника и анального отверстия . Вероятно, дикинсонии питались посредством поглощения питательных веществ нижней поверхностью тела и, таким образом, они занимают промежуточное положения между губками и эуметазоями и, возможно, родственны пластинчатым .
Кладограмма
Кладограмма взаимоотношений между Dipleurozoa (Dzik, 2003 )
|
|||||||||||||||||||||
См. также
Примечания
- Малаховская Я. Е., Иванцов А. Ю. Двусторонне-симметричные поперечно-расчлененные формы // . — Архангельск : Издательство ПИН РАН, 2003. — 48 с.
- Еськов К. Ю. Словарь названий живых и ископаемых организмов // Удивительная палеонтология: история Земли и жизни на ней. — М. : ЭНАС, 2008. — 312 с. — ISBN 978-5-91921-129-7 .
- Иванцов А. Ю., Леонов М. В. Отпечатки вендских животных — уникальные палеонтологические объекты Архангельской области. — Архангельск: Дирекция ООПТ, 2008. — С. 26—31. — 96 с.
- Крылов Н. Н. Органический мир докембрия // Розанов А. Ю. , Миссаржевский В. В., Волкова Н. А. и др. от 9 января 2018 на Wayback Machine (Труды ГИН АН СССР . Вып. 206) / ред. кол. акад. А. В. Пейве (гл. ред.), К. И. Кузнецова, акад. В. В. Меннер , П. П. Тимофеев , отв. ред. М. Е. Раабен. — М.: Наука, 1969. — С. 261. — 380 с.
- Крылов И. Н. от 27 декабря 2017 на Wayback Machine / отв. ред. акад. В. В. Меннер . — М.: Наука, 1972. — С. 89. — 105 с.
- ↑ Bobrovskiy Ilya , Hope Janet M. , Ivantsov Andrey , Nettersheim Benjamin J. , Hallmann Christian , Brocks Jochen J. (англ.) // Science. — 2018. — 20 September ( vol. 361 , no. 6408 ). — P. 1246—1249 . — ISSN . — doi : .
- Марков А. // Элементы.ру . — 2018. — 24 сентября.
- Ivantsov, A. Yu. Small Vendian transversely Articulated fossils // Paleontological Journal . — Nauka , 2007. — Т. 41 , № 2 . — С. 113—122 . — doi : .
- B. M. Keller and M. A. Fedonkin. (рус.) // Izv. Akad. Nauk SSSR, Ser. Geol.. — 1976. — Т. 3 . — С. 38—44 . 27 сентября 2007 года. . Дата обращения: 22 октября 2011. Архивировано 27 сентября 2007 года.
- Evans, S.D.; Hunt, G.; Gehling, J.G.; Sperling, E. A.; Droser, M.L. (2023). "Species of Dickinsonia Sprigg from the Ediacaran of South Australia". Palaeontology (e12635): 1—21. doi : .
- ↑ Retallack, G.J. (англ.) // Alcheringa: an Australasian Journal of Palaeontology : journal. — 2007. — Vol. 31 , no. 3 . — P. 215—240 . — doi : .
- ↑ Малаховская Я. Е., Иванцов А. Ю. . Химия и жизнь, 2004, № 7.
- Seilacher 1989
- Dzik 2000??
- Sprigg, Reg C. (англ.) // Trans. Roy. Soc. S. Aust. : journal. — 1947. — Vol. 71 . — P. 212—224 . 29 сентября 2007 года. . Дата обращения: 22 октября 2011. Архивировано 29 сентября 2007 года.
- Retallack, G. J., Matthews, N. A., Master, S., Khangar, R. G., & Khan, M. Dickinsonia discovered in India and late Ediacaran biogeography (англ.) // Gondwana Research : журнал. — 2021. — Vol. 90 . — P. 65—170 .
- Wang, X. P., Chen, Z., Pang, K., Zhou, C. M., Xiao, S., Wan, B., & Yuan, X. L. Dickinsonia from the Ediacaran Dengying Formation in the Yangtze Gorges area, South China (англ.) // Palaeoworld : журнал. — 2021. — Vol. 30 , no. 4 . — P. 602-609 . — ISSN .
- Grazhdankin, Dima. (англ.) // Palæobiology : journal. — 2004. — Vol. 30 , no. 2 . — P. 203—221 . — doi : . 26 июля 2011 года.
- Александр Марков. . Элементы.ру (24 сентября 2018). Дата обращения: 12 декабря 2019. 14 декабря 2019 года.
- Ivantsov A. Y., Malakhovskaya Y. E. (неопр.) // Doklady Earth Sciences. — 2002. — Т. 385 , № 6 . — С. 618—622 . 4 июля 2007 года. . Дата обращения: 22 октября 2011. Архивировано из 4 июля 2007 года.
- Jensen, Sören; Droser, Mary L.; Gehling, James G. Trace fossil preservation and the early evolution of animals (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology : journal. — 2005. — Vol. 220 , no. 1—2 . — P. 19—29 . — doi : .
- Ivantsov, A. Yu. Feeding traces of Proarticulata — the Vendian metazoa (англ.) // Paleontological Journal : journal. — Nauka , 2011. — Vol. 45 , no. 3 . — P. 237—248 . — doi : .
- Ivantsov, A. Yu. (англ.) // Paleontological Journal : journal. — Nauka , 2004. — Vol. 38 , no. 3 . — P. 247—253 . 27 сентября 2007 года. . Дата обращения: 22 октября 2011. Архивировано 27 сентября 2007 года.
- ↑ Retallack, G.J. Were the Ediacaran fossils lichens? (англ.) // . — , 1994. — Vol. 17 . — P. 523—544 . — ISSN .
- Retallack, Gregory J. (2004) "Death, Decay and Destruction of Dickinsonia ". Geological Society of America Abstracts with Programs , Vol. 36, No. 5, p. 521 от 6 марта 2016 на Wayback Machine
- Dzik, Jerzy. (2000) "The Origin of the Mineral Skeleton in Chordates." in Max Knobler Hecht, Ross J. MacIntyre and Michael T. Clegg, eds. Evolutionary Biology Vol. 31. Pp. 105—146. Springer. ISBN 0-306-46178-1 от 10 ноября 2012 на Wayback Machine – URL retrieved February 10, 2007
- Waggoner, B.; Collins, A.G. Reductio Ad Absurdum : Testing The Evolutionary Relationships Of Ediacaran And Paleozoic Problematic Fossils Using Molecular Divergence Dates (англ.) // : journal. — , 2004. — Vol. 78 , no. 1 . — P. 51—61 . — ISSN . — doi : .
- Runnegar Bruce. (англ.) // Alcheringa: An Australasian Journal of Palaeontology. — 1982. — Т. 6 . — С. 223—239 . — doi : . 25 мая 2021 года.
- ↑ от 22 ноября 2011 на Wayback Machine — на сайте medbiol.ru.
- Brasier, M.; Antcliffe, J. Decoding the Ediacaran Enigma (англ.) // Science. — 2004. — Vol. 305 , no. 5687 . — P. 1115—1117 . — doi : . — .
- ↑ Sperling, Erik; Vinther, Jakob; Pisani, Davide; Peterson, Kevin (2008). 15 февраля 2010 года. . В кн.: Cusack, M; Owen, A; Clark, N. Programme with Abstracts. 52 . от 5 ноября 2013 на Wayback Machine . Glasgow, UK. p. 81.
- Sperling, Erik; et al. (англ.) // Geological Society of America Abstracts with Programs : journal. — 2008. — Vol. 40 , no. 6 . — P. 508 . 28 февраля 2018 года.
- Dzik J. (2003). "Anatomical Information Content in the Ediacaran Fossils and Their Possible Zoological Affinities" (pdf). Integrative and Comparative Biology. 43 (1): 114–126.
| Проартикуляты | |
|---|---|
| Петалонамы | |
| Трилобозои | |
|
Предположительно
современные типы |
|
|
Неопределённого
положения |
|
- 2021-12-18
- 1