Interested Article - Эндоспоры

- 2021-04-14
- 1

Эндоспо́ры — покоящиеся формы бактерий , которые образуются в результате скоординированной дифференцировки двух дочерних клеток , образовавшихся при удвоении генома исходной бактерии, причём одна из них проникает внутрь цитоплазмы другой дочерней клетки, которая становится внешней. Далее внешняя дочерняя клетка (её иногда называют спорангием ) погибает программируемой гибелью , а внутренняя клетка (преспора) становится собственно эндоспорой и входит в состояние максимального физиологического покоя, при котором все физиологические процессы внутри споры останавливаются ( гиперанабиоза ). Она становится чрезвычайно резистентной к неблагоприятным условиям окружающей среды и может сохранять жизнеспособность в течение длительного времени .
Описаны два случая, когда материнская клетка даёт не одну, а две эндоспоры. Так, крупная (35 мкм длиной) некультивируемая бактерия , обитающая в желудочно-кишечном тракте некоторых грызунов , образует до девяти эндоспор на спорангий. Другая гигантская некультивируемая бактерия, sp. (она может достигать размера 60 × 300 мкм), обитающая внутри тропической рыбы -единорога , образует две эндоспоры в одном спорангии .
Спорогенные бактерии
Эндоспоры образуют представители около 25 родов из типа фирмикут : Bacillus , Clostridium , , , , , , , , , , , , , , , и другие. В подавляющем большинстве случаев спорогенные бактерии являются грамположительными , хотя есть и исключения. Например, и относятся к числу грамотрицательных бактерий .
Строение эндоспоры

Основные структурные элементы зрелой эндоспоры:
- цитоплазматический компартмент, или кор;
- комбинированная клеточная стенка , состоящая из внутренней стенки и наружной стенки (кортекса);
- цитоплазматическая мембрана эндоспоры (внутренняя мембрана );
- цитоплазматическая мембрана спорангия (внешняя мембрана);
- внутренний и внешний споровые покровы;
- наружный покров ( экзоспорий ) .

Расположение эндоспор внутри материнской клетки может быть различным, и тип спорообразования в некоторых случаях используют при идентификации неизвестной культуры. Иногда формирующаяся эндоспора достигает таких размеров, что расширяет спорангий посередине или с одного из концов .
Кор эндоспоры содержит хромосому и небольшие количества компонентов аппарата трансляции : рибосомы , тРНК , сопутствующие ферменты и белковые факторы. При этом нестабильные компоненты клеток, такие как мРНК и нуклеозидтрифосфаты , отсутствуют, но есть запасы стабильных предшественников АТФ — АДФ и АМФ .
Сложная система покровов эндоспоры делает её высокорезистентной к различным неблагоприятным факторам. Её белковые покровы перекрывают доступ к кортексу ферментам , разрушающим пептидогликан (например, лизоциму ), а также защищают спору от агрессивных химических реагентов, таких как хлороформ . Мембраны спорангия и споры создают барьер проницаемости вокруг кора, не давая пройти к нему гидрофильным соединениям массой более 300 Да . Кроме того, низкое содержание воды в коре препятствует депуринизации геномной ДНК . Катионы двухвалентных металлов (прежде всего кальция ) хелатируются молекулами , которые синтезируются в спорангии, а потом доставляются в кор, что обеспечивает дополнительную защиту для ДНК. В коре эндоспор некоторых бактерий имеются особые белки, связывающиеся с ДНК и защищающие её не только от депуринизации, но и от разрушения сахарофосфатного остова. Наконец, прорастание эндоспоры сопровождается интенсивной репарацией повреждений в ДНК, накопленных во время периода покоя .
Споруляция
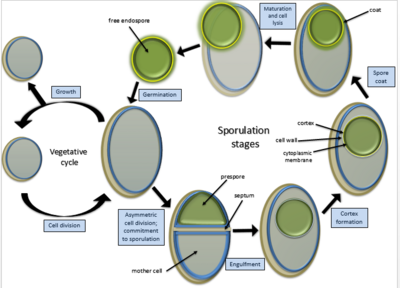
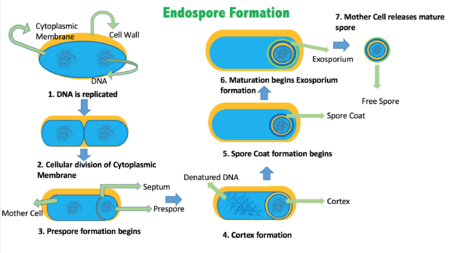
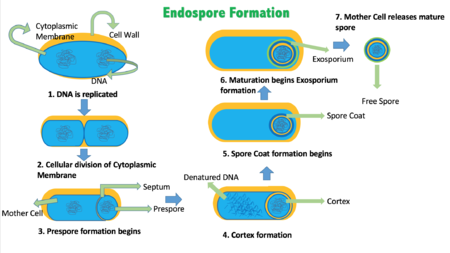
Цикл спорообразования протекает гораздо дольше (около 7 ч при 37 °С), чем деление с образованием других специализированных клеток. У Bacillus subtilis цикл споруляции включает семь стадий (0—VII), которые были предложены в 1960-х годах французским учёным Антуанетом Ритером.
- Стадия 0 . Исходная вегетативная клетка.
- Стадия I . В настоящее время эту стадию не выделяют, так как характерных морфологических признаков она лишена, а мутанты , не способные её проходить, неизвестны.
- Стадия II подразделяется на три этапа. На этапе IIi происходит септирование: в материнской клетке образуется споровая септа и происходит асимметричное бинарное деление , при нём образуются протопласты разного размера. На этапах IIii—IIiii больший протопласт начинает поглощать меньший.
- Стадия III . Поглощение завершается, и меньший протопласт (преспора) образуется в цитоплазме большей клетки (спорангия).
- Стадия IV . Происходит синтез модифицированной клеточной стенки — кортекса, который окружает преспору.
- Стадия V . В цитоплазме спорангия откладываются белковые покровы будущей эндоспоры.
- Стадия VI . Стадия морфологически не выражена, во время неё происходит созревание эндоспоры. Она приобретает повышенную резистентность к факторам окружающей среды, входит в гиперанабиоз, становится способной к прорастанию.
- Стадия VII . Спорангий подвергается программируемой гибели и частично лизируется , из-за чего зрелая эндоспора выходит наружу .
Уникальная споровая септа закладывается при симметричном образовании двух Z-колец на ¼ и ¾ материнской клетки. Одно из них случайным образом выбирается при участии белка SpoIIA, второе разрушается. Далее в области Z-кольца, как при обычном делении , происходит кольцевая инвагинация мембраны, впоследствии заполняющаяся пептидогликаном . Далее он лизируется, и в результате сестринские компартменты оказываются разделёнными только двумя мембранами. Хромосома будущей эндоспоры, по последним данным, проникает в меньший компартмент после смыкания септы. Её перенос опосредует ДНК- транслоказа FtsK/SpoIIIE, формирующая в мембранах компартментов два сквозных кольцевых гексамерных канала, и C-концевые домены канальцевых белков снабжают процесс энергией АТФ . Процесс поглощения меньшего компартмента большим в целом соответствует эндоцитозу (который вообще бактериям не свойственен), его молекулярные механизмы изучены слабо .
В лабораторных условиях бактерии приступают к споруляции, когда в среде заканчивается легко метаболизируемый источник углерода , азота или фосфора , однако триггеры споруляции в естественной среде обитания бактерий неизвестны. Ключевое событие начала споруляции — фосфорилирование транскрипционного фактора Spo0A с помощью протеинкиназ . Фосфорилирование Spo0A может запускаться тремя путями:
- Через киназу KinA, которая регистрирует окислительно-восстановительный и энергетический статус клетки.
- Через снижение уровня ГДФ и ГТФ в клетке, что свидетельствует о нехватке питательных веществ .
- Через белки, связанные с чувством кворума , то есть при достижении определённой плотности популяции бактерий .
В условиях, не благоприятствующих споруляции, фосфорилированный Spo0A дефосфорилируется фосфатазой Spo0E. Активный фосфорилированный Spo0A активирует транскрипцию регуляторов споруляции SpoIIA, SpoIIE и SpoIIG. Клетки, содержащие фосфорилированный Spo0A, выделяют белки, убивающие соседние клетки с неактивным Spo0A, чтобы использовать при споруляции их ресурсы. Также в запуске споруляции играет роль сигма-фактор σ H , контролирующий гены spoIIID и spoIIIM , их продукты участвуют в образовании споровой септы. После формирования септы активность Spo0A возрастает в спорангии, но снижается в преспоре. Далее в преспоре активируется сигма-фактор σ F , а в спорангии — σ E . Также в преспоре синтезируется σ G , который активируется только при завершении поглощения, а в спорангии на финальных стадиях спорообразования синтезируется сигма-фактор σ K . Совместно сигма-факторы преспоры и спорангия активируют экспрессию генов , необходимых для формирования эндоспоры .
Состояние покоя и прорастание
Находящаяся в состоянии покоя эндоспора характеризуется гиперанабиозом и гиперрезистентностью. Она не проявляет метаболической активности, не содержит важнейших метаболитов , таких как АТФ и ацетил-CoA , находящиеся в ней ферменты неактивны. В состоянии гиперанабиоза споры могут сохранять жизнеспособность на протяжении огромного периода времени. Так, эндоспоры сибирской язвы в скотомогильниках сохраняют жизнеспособность в течение 500 лет, споры актиномицетов — до 7500 лет . Имеются сведения, что споры Bacillus sp. , добытые из кристаллов поваренной соли в Нью-Мексико , сохраняли жизнеспособность на протяжении 250 млн лет. Эндоспоры не погибают под действием высокой и низкой температуры , при высушивании, большом гидростатическом давлении , при УФ - и γ-излучении , под действием сильных окислителей , при повышенной кислотности и других неблагоприятных условиях . Споры некоторых бактерий выдерживают даже кипячение в течение часа и более, поэтому растворы и инструменты стерилизуют в автоклавах с температурами стерилизации до 121 °C .
При благоприятных условиях эндоспора прорастает, то есть выходит из спорангия и превращается в нормальную вегетативную клетку. Индукторы прорастания могут быть как физиологическими (некоторые аминокислоты и сахара , пуриновые нуклеозиды и другие соединения или их смеси), так и нефизиологическими (минеральные соли , экзогенный дипиколинат кальция, лизоцим, катионные детергенты , сублетальный , давление от 100 до 600 МПа ). Прорастание начинается через секунды после воздействия индуктора и далее от него не зависит. Сначала из эндоспоры выходят протоны , ионы калия , натрия и цинка , дипиколиновая кислота с хелатированными ионами Ca 2+ , входит вода. Далее с помощью специальных ферментов лизируется пептидогликан кортекса, продолжается регидратация, при которой кор набухает и стенка растягивается. Наконец возобновляется нормальный метаболизм и биосинтез макромолекул .
Визуализация
Эндоспоры хорошо просматриваются при помощи как светового , так и электронного микроскопа . Так как эндоспоры непроницаемы для многих красителей , их визуализируют как неокрашенные тельца на фоне остального прокрашенного содержимого бактериальной клетки. Существуют, однако, методы , с помощью которых споры становятся видны в световой микроскоп как синие тельца в розовой цитоплазме .
Примечания
- , с. 257.
- , с. 258—259.
- , с. 258.
- , с. 257—258.
- , с. 67—68.
- , с. 68—69.
- , с. 261—262.
- , с. 263.
- , с. 263—264.
- , с. 266—267.
- , с. 267—268.
- ↑ , с. 67.
- , с. 259—261.
- , с. 268—269.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб. : Издательство С.-Петербургского университета, 2009. — Т. III. — 457 с. — ISBN 978-5-288-04894-4 .
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп. — М. : Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0 .
- Современная микробиология / Под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля. — М. : Мир, 2005. — Т. 1. — 654 с.
| Патогенные бактерии | |
|---|---|
|
Микрофлора
человека |
|
|
Субстратная
специфичность |
|
| Дыхание | |
|
Генетика и
размножение |
|
| Покоящиеся формы | |
| См. также | |
- 2021-04-14
- 1