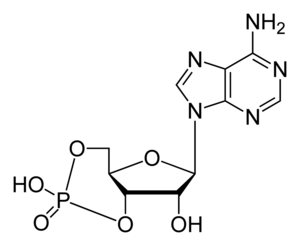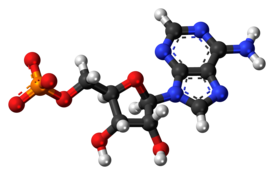
Аденозинмонофосфат
- 1 year ago
- 0
- 0



Циклический аденозинмонофосфат ( циклический AMФ , 3'5'-цAMФ , 3'5'-cAMP ) — органическое соединение , производное АТФ , выполняющее в организме роль вторичного посредника , использующегося для внутриклеточного распространения сигналов некоторых гормонов (например, глюкагона или адреналина ), которые не могут проходить через клеточную мембрану .
3'5'-цAMФ синтезируется аденилатциклазой в ответ на некоторые гормональные стимуляторы; действует как вторичный посредник при клеточном гормональном контроле путём стимуляции протеинкиназ . цАМФ является аллостерическим эффектором протеинкиназ A и ионных каналов. Синтезируется цАМФ мембранными аденилатциклазами (семейство ферментов, катализирующих реакцию циклизации АТФ с образованием цАМФ и неорганического пирофосфата ). Расщепление цАМФ с образованием АМФ катализируется фосфодиэстеразами . Ингибируются цАМФ только при высоких концентрациях метилированных производных ксантина , например, кофеина. Аденилатциклазы активируются G-белками (активность которых в свою очередь зависит от метаботропных рецепторов , связанных с G-белками) .
В неактивном состоянии протеинкиназа A является тетрамером , в котором две К (каталитические) субъединицы самоингибированы регуляторными (R) субъединицами. При связывании цAMФ R-субъединицы диссоциируют из комплекса и происходит активация К-субъединиц. Активированная протеинкиназа А фосфорилирует остатки серина и треонина в более чем 100 различных белках, в том числе во многих ферментах.
цAMФ осуществляет функции вторичного внутриклеточного посредника в действии первичных посредников (веществ, имеющих короткий период биодеградации) — например, ряда гормонов и нейромедиаторов. цAMФ опосредует биологическую функцию гормонов путём активации (инактивации) клеточных протеинкиназ (фосфатаз). Протеинкиназы, в свою очередь, фосфорилируют эффекторные белки и изменяют (увеличивают или уменьшают) их активность.
При активации аденилатциклазы , катализирующей образование цAMФ из АТФ, или блокировании фосфодиэстеразы , осуществляющей деградацию этого цAMФ, концентрация цAMФ в клетке увеличивается. Таким образом, содержание цАМФ в клетке определяется соотношением активностей этих двух ферментов. Связь между гормоном или др. химическим сигналом (первый посредник) и цAMФ (второй посредник) осуществляет аденилатциклазный комплекс, включающий рецептор, настроенный на определённый гормон (или др. биологически активное вещество) расположенный на внешней стороне клеточной мембраны, и аденилатциклазу, расположенную на внутренней стороне мембраны. Гормон, взаимодействуя с рецептором, активирует аденилатциклазу, которая образует цAMФ из АТФ.
Концентрация цAMФ, образующегося в клетке, превышает концентрацию действующего на клетку гормона в 100 раз. В основе механизма действия цAMФ в тканях животных и человека лежит его взаимодействие с протеинкиназами, например, протеинкиназы А. Связывание цAMФ с регуляторной субъединицей протеинкиназы приводит к диссоциации фермента и активации его каталитической субъединицы, которая, освободившись от регуляторной субъединицы, способна фосфорилировать определённые белки (в том числе ферменты). Изменение свойств этих макромолекул путём фосфорилирования меняет и соответствующие функции клеток. цAMФ играет определённую роль в морфологии, подвижности, пигментации клеток, в кроветворении, клеточном иммунитете, вирусной инфекции и др.
В бактериях уровень цАМФ изменяется в зависимости от среды культивирования. В частности, уровень цАМФ низок, если в качестве источника углерода используется глюкоза. Это регулируется через ингибирование цАМФ-образующего фермента, аденилатциклазы, как побочный продукт транспорта глюкозы в клетку. Транскрипционный фактор CRP (cAMP receptor protein), также называемый CAP (активатный белок генов катаболизма) формирует комплекс с цАМФ и таким образом становится возможным его связывание с ДНК. Комплекс CRP-цАМФ увеличивает экспрессию большого количества генов, включая некоторые ферменты, ответственные за запасание энергии независимо от глюкозы.
цАМФ, к примеру, вовлечен в положительную регуляцию lac оперона. В среде с низкой концентрацией глюкозы, цАМФ накапливается и связывается с аллостерическим сайтом транскрипционного регулятора CRP. Этот белок переходит в активную форму и связывается со специфическим сайтом левее lac промотора, облегчая посадку РНК полимеразы на соседний промотор для старта транскрипции с lac оперона, увеличивая скорость транскрипции lac оперона. При высокой концентрации глюкозы, концентрация цАМФ падает, и CRP диссоциирует из lac оперона.
|
|
|
|---|
|
Типы
нуклеиновых кислот
|
||||
|---|---|---|---|---|
| Азотистые основания | ||||
| Нуклеозиды | ||||
| Нуклеотиды | ||||
|
РНК
|
||||
|
ДНК
|
||||
| Аналоги | ||||
| Типы векторов | ||||
|
||||