Interested Article - Пелагические рыбы

- 2020-01-14
- 1

Пелагические рыбы — рыбы , обитающие в пелагической зоне океанов или озёр , в отличие от донных , которые живут на дне или у дна, и рифовых , связанных с прибрежной зоной .
Морские пелагические воды — крупнейшая акватория на Земле , объём которой по разным оценкам составляет 1370 миллионов кубических километров . Она служит средой обитания для более 11 % известных ныне видов рыб . Средняя глубина океанов составляет 4000 м. Свыше 98 % объёма воды находится глубже 100 м, а более 75 % глубже 1000 м .
Морских пелагических рыб можно разделить на океанических и прибрежных . Прибрежные рыбы населяют мелководье над континентальным шельфом , куда проникает солнечный свет. Океанические обитают в более глубоких и обширных водах за континентальным шельфом, хотя иногда заплывают в прибрежную зону .
Размер пелагических рыб колеблется от мелких , таких как сельди и сардины , до крупных хищников , занимающих вершину пищевой пирамиды , например, тунцов и океанических акул .
Как правило пелагические рыбы — отличные пловцы, они стремительно скользят в плотных слоях воды. У многих тело имеет веретенообразную или торпедообразную форму, позволяющую им развивать в воде высокую скорость и преодолевать большие расстояния . Парусник способен развить скорость до 109 км/ч , а некоторые виды тунцов пересекают Тихий океан . Многие пелагические рыбы образуют гигантские косяки массой свыше тысячи тонн. Другие, подобно рыбе-луне , чья масса достигает 2 тонн, ведут одиночный образ жизни .
У пелагических рыб плотное тело, они имеют отрицательную плавучесть . Почти у всех видов за редким исключением имеется плавательный пузырь, заполненный газовой смесью низкой плотности. Благодаря пузырю тело пелагических рыб обретает нейтральную или положительную плавучесть. В Мировом океане рыбы с плавательным пузырём встречаются на глубине до 7000 м. У пелагических рыб плавательный пузырь выполняет также гидростатическую функцию .

Экологические типы
Пелагических рыб можно разделить на два типа. Первый — активные хищники, которые охотятся на стайных или одиночных рыб среднего или небольшого размера. Обычно они держатся поодиночке или небольшими группами, собираясь в стаи при обнаружении добычи. У них хорошо развиты органы чувств и довольно сложное поведение. Типичные представители этого типа: синяя акула , мако , тунцы , меч-рыба , парусник . Второй тип — пастбищники-номады, которые совершают миграции и питаются планктоном и мелкими пелагическими рыбами. Это гигантская акула , манты и рыба-луна .

Эпипелагические рыбы
Эпипелагические рыбы обитают в эпипелагической зоне. Это верхний слой воды моря до глубины 200 м (нижняя граница сублиторали) . Также упоминается как поверхностные воды и включает в себя эвфотическую зону . Нижняя граница эвфотической зоны проходит на глубине, на которую проникает 1 % солнечного света, и где освещённость составляет 400 лк . Эта глубина зависит от мутности воды, но в прозрачной воде простирается до 200 метров и совпадает с границами эпипелагиали. В эвфотической зоне достаточно света для жизни фитопланктона и протекания фотосинтеза .

Обширная эпипелагическая зона служит средой обитания многих пелагических рыб. Она хорошо освещена, так что хищники могут охотиться с помощью зрения . Вода в ней активно перемешивается волнами и насыщена кислородом , поэтому в ней хорошо растут водоросли . Однако это довольно пустынное пространство. Однообразие условий среды обитания обуславливает отсутствие видового разнообразия — в этой зоне обитает всего 2 % известных видов рыб. На большей части эпипелагиали для рыб слишком мало корма, поэтому эпипелагических рыб чаще можно встретить в прибрежных водах над континентальным шельфом, куда либо апвеллинг приносят питательные вещества .
Эпипелагические рыбы делятся на мелких кормовых и крупных хищников, которые охотятся на них. Кормовые рыбы образуют косяки и планктон . У большинства видов тело вытянутое и обтекаемое, что позволяет развивать высокую скорость и совершать длительные миграции. В целом кормовые рыбы и хищники имеют схожее морфологическое строение. У хищников как правило веретенообразное гладкое тело, серповидный хвост и большой рот. Дорсальная поверхность тела эпипелагических рыб обычно окрашена в чёрный, синий или зелёный цвет, а бока и брюшко серебристые. Тёмная спина делает рыбу малозаметной на фоне дна. Серебристые светлые бока и брюшко плохо видны снизу на фоне зеркальной поверхности воды. Киль на брюхе уменьшает тень, образующуюся с брюшной стороны, которая может демаскировать рыбу. При взгляде на сбоку свет, падающий на тёмную спинку, и тень нижней части рыбы, скрадываемая блеском чешуи, делает рыбу незаметной . Свет всегда падает в воду сверху, освещая лучше всего спинку рыбы, брюшко, остаётся в тени. Освещённая тёмная спинка кажется светлее, чем есть, серые боках неизменны, а брюшко выглядит темнее. При взгляде сбоку рыба кажется сероватой и сливается с толщей воды. Кроме того, однотонная окраска без теней лишает предмет рельефности, рыба кажется плоской, её очертания теряют чёткость . Кожа эпипелагических рыб, как правило, покрыта блестящей чешуёй , каждая чешуйка служит своеобразным зеркалом . Свет отражается от вертикально ориентированной чешуи, делая рыбу на средней глубине почти невидимой при взгляде сбоку .


Несмотря на ограниченное количество видов, эпипелагические рыбы очень многочисленны. Отсутствие разнообразия восполняется числом. Кормовые рыбы образуют огромные косяки, а охотящиеся на них хищные рыбы являются в свою очередь ценными промысловыми объектами . Многие кормовые рыбы являются факультативными хищниками, которые при случае подхватывают из водяной толщи рачков или личинок, и возвращаются к фильтрации фитопланктона, когда это становится энергетически более эффективно. Фильтрация, как правило, осуществляется длинными жаберными тычинками . Некоторые крупнейшие эпипелагические рыбы, например, китовая и гигантская акула , так же как и самые мелкие шпроты и анчоусы, являются фильтраторами .
В совершенно прозрачных океанских водах мало корма. Области с его повышенным содержанием, как правило, мутные от цветущего планктона . К ним устремляются животные-фильтраторы, которые, в свою очередь, привлекают хищников. Лов тунца оптимально проводить, когда мутность воды, оценённая с помощью диска Секки на максимальной глубине в солнечный день, составляет 15—35 м .
Плавучие объекты
Эпипелагических рыб привлекают плавучие объекты. Они собираются в многочисленные стаи вокруг дрейфующих обломков, плотов, медуз и водорослей, «обеспечивающих в оптической пустоте визуальный стимул» . Плавучие объекты предоставляют некоторую защиту от хищников. У некоторых видов присутствие большого количества дрейфующих водорослей или медуз существенно увеличивает уровень выживаемости молодняка .

Молодь многих прибрежных рыб использует морские водоросли как укрытие и источник пищи, поскольку в них прячутся беспозвоночные и другие мелкие рыбы. Дрейфующие водоросли, в особенности саргасс , обеспечивают среду обитания с полноценным укрытием и кормовой базой и даже поддерживают собственную уникальную фауну , например, саргассового морского клоуна . В ходе исследования у побережья Флориды в одном скоплении саргассовых водорослей было обнаружено 54 вида животных из 23 семейств . Молодь рыб использует в качестве укрытия и источника пищи медуз, несмотря на то, что последние охотятся на мелких рыб . Чтобы не гонятся по всему океану за подвижными стаями пелагических рыб, например, тунцов, используют специальные . Они представляют собой заякоренные плоты или другие предметы, плавающие на или у поверхности воды. В Тихом и Индийском океанах рыбаки устанавливают плавучие манки из разного мусора вокруг тропических островов и кошельковыми неводами ловят привлечённую ими рыбу .

Исследования, проведённые с помощью эхолота во Французской Полинезии , обнаружили вокруг таких устройств крупные стаи желтопёрых и большеглазых тунцов , собравшиеся на расстоянии от 10 до 50 м. На большем удалении, от 50 до 150 м, наблюдались скопления желтопёрых тунцов и длиннопёрых тунцов меньшей плотности. Ещё дальше, на расстоянии свыше 500 м, были разбросанные группы крупных взрослых тунцов разных видов. Эти группы отличались по распределению и плотности и в некоторых местах перекрывали друг друга. Устройства привлекали и другие виды рыб. С наступление темноты группы рассеялись .
Крупных пелагических рыб, даже хищных, часто сопровождает свита из мелких рыбёшек. Фридайверы, которые долгое время остаются под водой, также могут привлечь к себе подобную свиту, при этом мелочь будет следовать по пятам, а крупные рыбы — наблюдать на расстоянии. Морские черепахи , которые служат естественным укрытием мелким рыбкам, иногда становятся невольной мишенью атаки марлинов , привлечённых их свитой .
Прибрежные рыбы

Рыб, обитающих в прибрежной зоне над континентальным шельфом, также называют неритическими . Поскольку глубина континентального шельфа обычно не превышает 200 м, прибрежные рыбы, которые не являются придонными, как правило, обитают в залитой солнцем эпипелагиали. Неритические эпипелагические рыбы — одни из самых распространённых рыб во всём мире. Среди них есть как кормовые, так и хищные. Кормовые рыбы процветают в прибрежных зонах, богатых питательными веществами за счёт апвеллинга и плоскостного стока. Некоторые виды являются проходными или полупроходными .

Океанические рыбы
Океанические рыбы живут в водах открытого моря за континентальным шельфом. Они могут сильно отличаться от неритических видов. Тем не менее, эти два типа не являются взаимоисключающими, поскольку между прибрежной и океанической областями нет чёткой границы и многие эпипелагические рыбы переходят из одной области в другую, особенно на различных этапах жизненного цикла .
Океанические эпипелагические рыбы бывают постоянными обитателями, частичными обитателями и случайными посетителями. Постоянные обитатели всю жизнь проводят в открытом море. К ним относятся лишь несколько видов, например, тунцы, летучие рыбы , марлины, макрелещуки (сайра), лоцманы , реморы , океанические акулы и обыкновенная луна-рыба . Большинство из этих видов бороздит открытой океан, изредка появляясь на континентальном шельфе. Перемещения некоторых постоянных обитателей связаны с дрейфом медуз или морских водорослей .
Частичные обитатели делятся на 3 группы: виды, у которых в этой зоне живёт только молодь; виды, обитающие в открытом море лишь во взрослом состоянии ( лососи , летучие рыбы, китовые акулы ) и глубоководные виды, совершающие суточные вертикальные миграции, которые ночью поднимаются к поверхности воды (например, миктофовые ) . Случайными посетителями океанической эпипелагиали бывают как взрослые особи, так и молодняк видов, постоянно живущих в других зонах и случайно заносимых сюда течением .
-
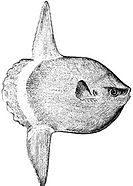 Обыкновенная луна-рыба , постоянный обитатель океанической эпипелагической зоны, иногда дрейфует с течением, поедая медуз
Обыкновенная луна-рыба , постоянный обитатель океанической эпипелагической зоны, иногда дрейфует с течением, поедая медуз -
 Китовая акула, ещё один обитатель океанической эпипелагиали, цедит планктон и периодически опускается в мезопелагическую зону
Китовая акула, ещё один обитатель океанической эпипелагиали, цедит планктон и периодически опускается в мезопелагическую зону -
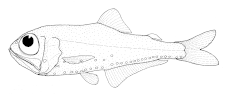 Светящийся анчоус является частичным обитателем океанической эпипелагической зоны. Днём эти рыбы держатся на глубине, а ночью в поисках пищи поднимаются к поверхности воды
Светящийся анчоус является частичным обитателем океанической эпипелагической зоны. Днём эти рыбы держатся на глубине, а ночью в поисках пищи поднимаются к поверхности воды
Глубоководные рыбы
Намного ниже эпипелагиали обитают различные типы пелагических рыб, приспособленные к жизни в этих более глубоких зонах .
В глубинах океана идёт непрерывный , самопроизвольное осаждение детрита из верхних слоёв водной толщи. Он является производным жизнедеятельности организмов продуктивной эвфотической зоны. Морской снег включает мёртвый или умирающий планктон, простейших ( диатомовые водоросли ), фекалии , песок , сажу и прочую неорганическую пыль. По пути «снежинки» растут и за нескольких недель, пока не опустятся на дно океана, могут достигать нескольких сантиметров в диаметре. Тем не менее, большинство органических компонентов морского снега потребляются микробами , зоопланктоном и другими животными-фильтраторами на протяжении первых 1000 метров их путешествия, то есть, в эпипелагиали. Таким образом, морской снег можно считать основой глубоководной мезопелагической и донной экосистем: поскольку солнечный свет не может проникнуть сквозь толщу воды, глубоководные организмы в качестве источника энергии полагаются на морской снег.
Некоторые группы глубоководных рыб, такие как представители семейств миктофовых, , топориковых , фотихтиевых , иногда называют псевдоокеаническими, поскольку они концентрируются вокруг структурных оазисов, в частности у подводных вершин и на материковом склоне. Подобное явление объясняется присутствием кормовой базы, которую в свою очередь привлекают подобные оазисы .
Рыбы различных пелагических и глубоководных донных зон заметно отличаются друг от друга по поведению и строению. Группы сосуществующих видов в пределах каждой зоны функционируют аналогичным образом, например, небольшие мезопелагические фильтраторы, совершающие вертикальные миграции , батипелагические удильщики и глубоководные донные долгохвостовые .
Среди видов, обитающих на глубине, редко встречаются . Вероятно, глубоководные рыбы — достаточно древние и настолько хорошо приспособлены к окружающей среде, что появление современных рыб не увенчалось успехом . Несколько глубоководных представителей колючепёрых принадлежат к древним отрядам бериксообразных и опахообразных . Большинство пелагических рыб, встречающихся на глубине, принадлежат к собственным отрядам, что предполагает длительную эволюцию в подобных условиях. И, напротив, глубоководные донные виды принадлежат к отрядам, которые включают многих рыб, обитающих на мелководье .
| Виды по зонам пелагиали | |
|---|---|
|
Многие виды совершают суточные вертикальные миграции между зонами.
В этой таблице они перечислены по средней или более глубокой зоне основного пребывания |
|
| Зона | Виды и группы видов: |
| Эпипелагическая |
|
| Мезопелагческая | светящиеся анчоусы , опахи , алепизавры , опистопроктовые , , меламфаевые, саблезубые |
| Батипелагическая | В основном гоностомовые и удильщики . А также длиннорогие саблезубы , хаулиоды , живоглоты , гигантуровые , , , , , циемовые , лофотовые , мешкоротообразные , китовидковые , топориковые |
| Бентопелагическая | Особенно многочисленны долгохвостовые и бититовые |
| Бентическая | Камбалы , зеленоглазковые , бельдюги , угри , скаты , пинагоры и лопаточные нетопыри |
| Сравнительное строение пелагических рыб | |||
|---|---|---|---|
| Параметр | Мезопелагические виды | Батипелагические виды | Глубоководные и бентические виды |
| мускулы | тело мускулистое | плохо развиты, тело дряблое | |
| скелет | крепкий | слабо окостеневший | |
| чешуя | да | нет | |
| нервная система | хорошо развита | только боковая линия и обоняние | |
| глаза | крупные и чувствительные | маленькие, иногда не функциональные | различные (от хорошо развитых до атрофированных) |
| фотофоры | часто имеются | часто имеются | обычно нет |
| жабры | хорошо развиты | ||
| почки | крупные | маленькие | |
| сердце | крупное | маленькое | |
| плавательный пузырь | имеется у рыб, совершающих вертикальные миграции | редуцирован или отсутствует | различный (от хорошо развитого до полного отсутствия) |
| размер | обычно менее 25 см | различный, виды длиной 1 м встречаются редко | |
Мезопелагические рыбы

Под эпипелагической зоной условия окружающей среды резко меняются. На глубине между 200 и 1000 м освещённость ослабевает до тех пор, пока не наступает полная темнота. Через термоклин температура падает до 4—8 °C. Это сумеречная или . Давление продолжает нарастать на 1 атмосферу каждые 10 м, тогда как концентрация пищи, содержание кислорода и скорость циркуляции воды снижаются .
Радисты, использующие разработанные во время Второй Мировой войны сонары , были озадачены, обнаружив днём на глубине 300—500 метров ложное морское дно. Ночью подобное явление наблюдалось на меньшей глубине. Оказалось, что оно связано с присутствием миллионов морских организмов, в особенности мелких мезопелагических рыб, обладающих плавательным пузырём, которые отражали сигнал гидролокатора. В сумерках рыбы поднимались к поверхности, чтобы кормиться планктоном. Этот феномен известен как звукорассеивающий слой . Слой опускается, когда на небе сияет луна , и поднимается, если она закрыта облаками .
Большинство мезопелагических рыб совершают суточные вертикальные миграции, поднимаясь ночью в эпипелагическую зону, зачастую вслед за перемещением планктона, и возвращаясь днём на безопасную глубину. Подобные вертикальные миграции часто имеют большую протяжённость, они осуществляется с помощью плавательного пузыря. Когда рыба хочет всплыть, пузырь надувается. Учитывая высокое давление в мезопелагической зоне, это требует значительной энергии. При подъёме давление в плавательном пузыре должно регулироваться, чтобы его не разорвало. Если рыба собирается вернуться на глубину, пузырь сдувается . В ходе миграций некоторые мезопелагические рыбы проходят через термоклин, где температура меняется с 10 до 20 °C, что говорит об их терпимости к быстрым температурным изменениям .

У этих рыб мускулистое тело, покрытое чешуёй, крепкий скелет, хорошо развитые жабры и центральная нервная система, крупные сердце и почки. У мезопелагических фильтраторов маленький рот и тонкие жаберные тычинки, тогда как у ихтиофагов рот крупный, а тычинки грубее . У рыб, совершающих вертикальные миграции имеется плавательный пузырь .
Мезопелагические рыбы приспособлены к активному образу жизни в условиях низкой освещённости. Большинство из них — хищники с крупными глазами. У некоторых глубоководных видов трубчатые глаза с большими линзами, которые глядят вверх, а из фоторецепторов присутствуют лишь палочки . Это обеспечивает бинокулярное зрение и высокую чувствительность к слабым световым сигналам . Такая адаптация обеспечивает повышенное конечное видение за счёт бокового видения, и позволяет хищникам высмотреть кальмаров, каракатиц и мелкую рыбу, силуэт которых вырисовывается над ними на фоне темноты.
Обычно мезопелагические рыбы лишены защитных шипов, их защитой служит камуфляж . Хищники окрашены в тёмные цвета, чёрный или красный. На глубине длинноволновый красный цвет невидим, и красная окраска столь же эффективна, как и чёрная. Рыбы, совершающие вертикальные миграции, окрашены контрастно в серебристых тонах, с тёмной спинкой и светлым брюшком. На вентральной поверхности тела у них зачастую имеются фотофоры, испускающие свет, маскирующий силуэт рыбы при взгляде снизу. Однако, глаза некоторых хищников оснащены жёлтыми линзами, фильтрующие рассеянный свет, которые делают видимым биолюминесцентное свечение .
-
![У антарктического клыкача крупные, обращённые вверх глаза, способные различить силуэт жертвы[33]](/images/005/101/5101538/16.jpg?rand=25040) У антарктического клыкача крупные, обращённые вверх глаза, способные различить силуэт жертвы
У антарктического клыкача крупные, обращённые вверх глаза, способные различить силуэт жертвы -
![У опистопроктовых глаза имеют форму бочонка, которые обычно направлены вверх, но могут нацелиться вперёд[34]](/images/005/101/5101538/17.jpg?rand=978579) У опистопроктовых глаза имеют форму бочонка, которые обычно направлены вверх, но могут нацелиться вперёд
У опистопроктовых глаза имеют форму бочонка, которые обычно направлены вверх, но могут нацелиться вперёд -
![У малакостов длина нижней челюсти равна 1/4 длины тела. Она лишена дна и крепится посредством шарнира и видоизменённой подъязычной кости. Позади крупных передних клыков расположены мелкие зазубренные зубы[35][36]](/images/005/101/5101538/18.jpg?rand=372840) У малакостов длина нижней челюсти равна 1/4 длины тела. Она лишена дна и крепится посредством шарнира и видоизменённой подъязычной кости . Позади крупных передних клыков расположены мелкие зазубренные зубы
У малакостов длина нижней челюсти равна 1/4 длины тела. Она лишена дна и крепится посредством шарнира и видоизменённой подъязычной кости . Позади крупных передних клыков расположены мелкие зазубренные зубы -
![Малакосты — одни из немногих рыб, способных испускать красный свет. Поскольку большинство жертв его не различает, хищник охотится с помощью невидимого луча[35]](/images/005/101/5101538/19.jpg?rand=712472) Малакосты — одни из немногих рыб, способных испускать красный свет. Поскольку большинство жертв его не различает, хищник охотится с помощью невидимого луча
Малакосты — одни из немногих рыб, способных испускать красный свет. Поскольку большинство жертв его не различает, хищник охотится с помощью невидимого луча
, принадлежащий семейству опистопроктовых, является единственным известным видом среди позвоночных , который фокусирует изображение с помощью зеркала , а не линзы .
Исследования, полученные с помощью учётного траления, показали, что светящиеся анчоусы формируют до 65 % биомассы всех глубоководных рыб . Действительно, миктофовые являются одними из наиболее распространённых по численности и разнообразных среди всех позвоночных, они играют важную экологическую роль в качестве пищи для крупных животных. Глобальная биомасса миктофовых оценивается в 660 млн тонн . Именно светящиеся анчоусы составляют большую часть биомассы, образующую глубинный рассеивающий слой мирового океана. Сигнал сонара отражается от миллионов плавательных пузырей этих рыб, создавая эффект ложного дна .
Большеглазые тунцы совершают вертикальные миграции, переходя из эпипелагической в мезопелагическую зону. Днём они опускаются на глубину до 500 м, вероятно, следуя за кормовыми рыбами .
-
![Алепизавры охотятся из засады. Это постоянные обитатели мезопелагической зоны. Один из самых крупных мезопелагических видов рыб, достигающий в длину 2 м[44]](/images/005/101/5101538/20.jpg?rand=320556) Алепизавры охотятся из засады. Это постоянные обитатели мезопелагической зоны. Один из самых крупных мезопелагических видов рыб, достигающий в длину 2 м
Алепизавры охотятся из засады. Это постоянные обитатели мезопелагической зоны. Один из самых крупных мезопелагических видов рыб, достигающий в длину 2 м -
![У представителей семейства гигантуровых крупные, направленные вперёд глаза с большими линзами[45]](/images/005/101/5101538/21.jpg?rand=762415) У представителей семейства гигантуровых крупные, направленные вперёд глаза с большими линзами
У представителей семейства гигантуровых крупные, направленные вперёд глаза с большими линзами -
![Кинжалозубыruen укусом обездвиживают прочих мезопелагических рыб[46]](/images/005/101/5101538/22.jpg?rand=604215) укусом обездвиживают прочих мезопелагических рыб
укусом обездвиживают прочих мезопелагических рыб -
 Большеглазые тунцы ночью бороздят воды эпипелагиали, а день проводят в мезопелагической зоне.
Большеглазые тунцы ночью бороздят воды эпипелагиали, а день проводят в мезопелагической зоне.
Батипелагические рыбы

соответствует открытой части океана над материковым склоном. Она расположена на глубине от 200—500 до 3000—4000 м . Это промежуточная глубинная зона между поверхностными зонами и . Она принадлежит к афотической зоне , которую иногда называют «ночной». Для неё характерно полное отсутствие солнечного света, огромное давление, низкая температура и малая концентрация растворённого в воде кислорода .
У батипелагических рыб имеются специальные адаптации , позволяющие им выжить в таких условиями — у них замедленный метаболизм и неспециализированная диета. Они предпочитают сидеть и ждать пищу, а не тратить энергию на её поиски. Поведение рыб батипелагиали можно противопоставить поведению мезопелагических рыб. Последние зачастую очень подвижны, в то время как многие батипелагические виды — это хищники, ожидающие в засаде, как правило, расходующие мало энергии на движение .
Среди батипелагических рыб преобладают небольшие удильщик гоностомовые. Также распространены длиннорогие саблезубы, рыбы-гадюки , кинжалозубы и веретенниковые. Это мелкие виды длиной около 10 см, и лишь немногие достигают более 25 см. Большую часть времени они почти неподвижно ожидают в толще воды появления добычи, привлекаемой светом их фотофор. Та немногая энергия, попадающая в батипелагическую зону, проникает сверху в виде детрита, фекалий и изредка беспозвоночных и мезопелагических рыб . Около 20 % пищи, произведённой в эпипелагиали, достигает мезопелагической зоны , тогда как в батипелагиали оседает лишь 5 % .

Батипелагические рыбы ведут малоподвижный образ жизни, они приспособлены к минимальному расходу энергии в естественной среде с очень скудной пищей и доступной энергии, где даже нет солнечного света, а есть лишь биолюминесценция. У них вытянутые тела дряблыми, водянистыми мышцами и слабый скелет. Поскольку они в основном состоят из воды, огромное давление на этих глубинах не оказывает на них сильного воздействия. У многих видов шарнирные челюсти, способные сильно распахиваться, оснащены загнутыми зубами. Кожа покрыта слизью и лишена чешуи. Центральная нервная система ограничена боковой линией и обонятельными структурами, глаза маленькие, зачастую нефункциональные, жабры, почки и сердце маленькие, плавательный пузырь, как правило, отсутствует .

Те же самые адаптации характерны и для личинок рыб батипелагиали. Это даёт основание предположить, подобные функции были приобретены в процессе эволюции через неотению . Даже на стадии личинки рыбы способны зависать в толще воды, затрачивая минимум энергии .
Несмотря на свирепую внешность, эти, как правило, миниатюрные рыбы со слабыми мышцами не представляют угрозу для человека. У глубоководных рыб плавательный пузырь либо совершенно отсутствуют, либо еле функционирует, и батипелагические рыбы обычно не совершают вертикальных миграций. Заполнение пузыря при таком большом давлении сопряжено с огромными энергозатратами. У некоторых глубоководных рыб плавательный пузырь функционирует на ранней стадии жизненного цикла, пока они обитают в верхней эпипелагической зоне, но с возрастом усыхает или заполняется жиром, когда рыба перемещается в среду обитания, характерную для взрослых особей .
Наиболее важными сенсорными системами, как правило, являются внутреннее ухо , воспринимающее звуки, и боковая линия , которая реагирует на перемену давления воды. Обонятельная система также может играть существенную роль для самцов, которые находят самок по запаху . Батипелагические рыбы окрашены в чёрный, реже в красный цвет. С помощью фотофоры они, как правило, приманивают добычу или привлекают сексуального партнёра. Поскольку еды очень мало, батипелагические хищники не отличаются разборчивостью, они хватают всё, что проплывает достаточно близко. Большой рот с острыми зубами служит для захвата крупной добычи, а перекрывающиеся жаберные тычинки не дают ускользнуть мелочи .
В этой зоне трудно найти партнёра для размножения. Некоторые виды зависят от биолюминесценции. Другие являются гермафродитами . Самки удильщиков вырабатывают феромоны , чтобы привлечь крошечных самцов. Найдя самку, самец кусает её и более никогда не отпускает. Самец после укуса выпускает фермент , который расщепляет кожу его рта и её тела, пара срастается, так что кровеносная система у них становится единой. Затем самец атрофируется, становясь не более чем парой половых желез . Такой экстремальный половой диморфизм гарантирует, что, когда самка готова к нересту, у неё в доступности будет источник спермы .
Помимо рыб в батипелагической зоне обитают кальмары , осьминоги , губки , брахиоподы , морские звёзды и морские ежи . Сюда опускаются крупные киты.
-
 Мешкоротообразные широко распахивают пасть и ловят ею, как неводом, добычу, которую привлекают люминесцентным органом на кончике хвоста.
Мешкоротообразные широко распахивают пасть и ловят ею, как неводом, добычу, которую привлекают люминесцентным органом на кончике хвоста. -
![Чёрный живоглот со своим растягивающимся желудком способен проглотить целиком добычу, которая весит больше него в 10 раз[58][59].](/images/005/101/5101538/28.jpg?rand=855548) Чёрный живоглот со своим растягивающимся желудком способен проглотить целиком добычу, которая весит больше него в 10 раз .
Чёрный живоглот со своим растягивающимся желудком способен проглотить целиком добычу, которая весит больше него в 10 раз . -
 Самец гаплофрины нежной прирастает к самке и атрофируется до пары гонад, становясь по мере необходимости источником спермы.
Самец гаплофрины нежной прирастает к самке и атрофируется до пары гонад, становясь по мере необходимости источником спермы. -
![У широкораспространённых саблезубов самые крупные среди рыб зубы относительно тела[60]. Несмотря на устрашающую внешность, батипелагические рыбы обычно слишком малы и слабы, чтобы представлять опасность для человека.](/images/005/101/5101538/30.jpg?rand=3050) У широкораспространённых саблезубов самые крупные среди рыб зубы относительно тела . Несмотря на устрашающую внешность, батипелагические рыбы обычно слишком малы и слабы, чтобы представлять опасность для человека.
У широкораспространённых саблезубов самые крупные среди рыб зубы относительно тела . Несмотря на устрашающую внешность, батипелагические рыбы обычно слишком малы и слабы, чтобы представлять опасность для человека. -
![Хаулиод обыкновенныйruen ночью может совершать вертикальные миграции, поднимаясь с батипелагических глубин почти на поверхность воды[61].](/images/005/101/5101538/31.jpg?rand=882913) ночью может совершать вертикальные миграции, поднимаясь с батипелагических глубин почти на поверхность воды .
ночью может совершать вертикальные миграции, поднимаясь с батипелагических глубин почти на поверхность воды .
Демерсальные рыбы

Демерсальные рыбы большую часть жизненного цикла проводят на дне или в непосредственной близости от дна (придонные рыбы) . Они встречаются как в прибрежных районах континентального шельфа, так и в открытом океане вдоль континентального склона. В целом они отсутствуют в абиссопелагической и ультраабиссальной зоне и на абиссальной равнине . Они занимают морское дно, покрытое илом , песком , гравием или валунами .
В глубоководной зоне донные рыбы довольно многочисленны и активны по сравнению с батипелагическими рыбами . Здесь распространены долгохвостовые , бититовые , бельдюги , лопаточные нетопыри и пинагоры .
У донных рыб хорошо развитые органы и мускулатура. По этим параметрам они ближе к мезопелагическим рыбам, чем к обитателям батипелагиали. Иными словами, они более разнообразны. У них как правило отсутствуют фотофоры. Глаза и плавательный пузырь могут быть как хорошо развитыми, так и атрофированными. Они сильно отличаются по размеру, нередко встречаются крупные экземпляры свыше 1 м длиной.
У многих демерсальных рыб длинное и узкое тело, подобное угрям. Возможно, это связано с длиной боковой линии, которая улавливает низкочастотные звуковые волны, тогда как некоторые рыбы с помощью мускулов издают подобные звуки, привлекая сексуальных партнёров . Обоняние также играет важную роль, судя по тому как быстро они ловятся на живца . Основу рациона демерсальных рыб составляют бентосные беспозвоночные и падаль . Они находят пищу в основном посредством боковой линии, обоняния и осязания .
Демерсальных рыб можно разделить на сугубо бентических и бентопелагических, имеющих отрицательную и нейтральную плавучесть соответственно. Бентические рыбы постоянно контактируют с дном. Они либо лежат в засаде в ожидании добычи, либо активно двигаются в поисках пищи .
Бентопелагические рыбы

Бентопелагические или придонные рыбы обитают в непосредственной близости у дна, питаются бентосом и бентопелагическим зоопланктоном . Большинство демерсальных рыб относятся к бентопелагическим . Их можно разделить на виды с крепким телом и дряблотелых. Дряблотелые бентопелагические виды схожи с батипелагическими, у них небольшая масса тела и низкий уровень метаболизма. Они затрачивают минимум энергии и охотятся из засады . Примером такого типа служит , хищник с крупной головой и телом, которое на 90 % состоит из воды. У этих рыб самые крупные глаза и самый маленький мозг относительно тела среди позвоночных .

Твёрдотелые бентопелагические рыбы — активные пловцы, которые энергично ищут добычу на дне. Иногда они живут вокруг подводных вершин с сильным течением . Примером такого типа являются патагонский клыкач и атлантический большеголов . Ранее эти рыбы водились в изобилии и являлись ценным объектом промысла, их добывали ради вкусного плотного мяса .
Бентические рыбы
Хотя бентические рыбы и не относятся к пелагическим, мы расскажем о них вкратце для полноты и сравнения.
Некоторые рыбе не подходят под представленную выше классификацию. Например, широко распространённые и почти слепые представители семейства , рацион которых состоит из бентопелагического зоопланктона. Тем не менее, они относятся к бентическим рыбам, поскольку находятся в постоянном контакте с дном. Они стоят на дне, опираясь на длинные лучи свои плавников головой к течению, и ловят проплывающую мимо пищу .
Самый глубоководный из известных ныне видов — Abyssobrotula galatheae , внешне похожие на угрей и совершенно слепые донные рыбы, которые питаются беспозвоночными .
-
![Батиптер Bathypterois grallatorruen «стоят» на дне, опираясь на длинные лучи своих плавников[73]](/images/005/101/5101538/35.jpg?rand=404445) «стоят» на дне, опираясь на длинные лучи своих плавников
«стоят» на дне, опираясь на длинные лучи своих плавников -
![Скат Taeniura meyeni охотятся на донных рыб, двустворчатых моллюсков, крабов и креветок[74]](/images/005/101/5101538/36.jpg?rand=634943) Скат Taeniura meyeni охотятся на донных рыб, двустворчатых моллюсков , крабов и креветок
Скат Taeniura meyeni охотятся на донных рыб, двустворчатых моллюсков , крабов и креветок

На больших глубинах, дефицит пищи и чрезвычайно высокое давление ограничивает выживаемость рыб. Самая глубокая точка океана находится на глубине около 11 000 метров. Батипелагические рыбы обычно не встречаются ниже 3000 метров. Наибольшая глубина обитания донных рыб составляет 8,370 м . Возможно, экстремальное давление подавляет важнейшие функции ферментов .
Бентические рыбы чаще попадаются и более разнообразны на континентальном склоне , где варьируется среда обитания и больше корма. Около 40 % дна океана состоит из абиссальных равнин , но эти плоские, пустынные регионы покрыты морскими отложениями и обычно придонная жизнь ( бентос ) здесь отсутствует. Глубоководные донные рыбы более распространены в каньонах или на скалах посреди равнин, где концентрируются сообщества беспозвоночных организмов. Подводные горы омываются глубоководными течениями, это становится причиной возникновения апвеллинга, который поддерживает жизнь донных рыб. Горные хребты могут делить подводные регионы на различные экосистемы .
Пелагический рыбный промысел
Кормовые рыбы

Крупные пелагические хищники обычно охотятся на мелких рыб пелагиали ( ). Кормовые рыбы кормятся, фильтруя планктон и, как правило, не превышают 10 сантиметров в длину. Они часто образуют косяки и мигрируют на большие расстояния между нерестилищами и местами нагула. Они особенно часто встречаются в апвеллинговых районах северной части Атлантического океана, у берегов Японии и у западного побережья Африки и Америки. У них, как правило, короткий срок жизни, и их заметно колеблются в течение года .
Атлантическая сельдь встречается в Северном море и Северной Атлантике на глубинах до 200 метров. Важный сельдевый промысел существовал в этих местах на протяжении веков. В прошлом в Норвегии основная часть зимнего улова жирной и молодой сельди приходилась на весенний нерест. Масштабная техническая модернизация рыбного промысла в 60-е годы привела к перелову — сильному истощению поголовья этого вида в начале 70-х годов. Было введено регулирование, и поголовье удалось восстановить .
Широкомасштабный промысел северной путассу начался в 1980-х годах, к моменту истощения запасов многих традиционных промысловых рыб. Сейчас объём вылова составляет до 2 млн тонн, российская квота составляет 60 тыс. т. Путассу ловят пелагическим тралом и кошельковым неводом . Важнейшим промысловым видом является сардина европейская . Эти стайные рыбы эпипелагиали обитают на глубине 25—50 м. Уровень запасов сардин достаточно велик, а промысел не наносит серьёзного вреда экосистемам. Их ловят жаберными сетями, кошельковыми неводами, тралами. В 1980-е годы вёлся массовый промысел (до 5 млн т. ежегодно) тихоокеанской сардины-иваси , но сейчас уловы существенно снизились — около 200 тыс. т. Изменения численности популяции этих рыб обусловлены климатическими факторами .
-
 Косяк перуанских сардин
Косяк перуанских сардин -
 Сельди кормятся рачками
Сельди кормятся рачками -
 Мойва — пелагическая кормовая рыба
Мойва — пелагическая кормовая рыба -

Хищные рыбы
См. также: Тунцы
К пелагическим рыбам среднего размера относятся летучие рыбы, ставриды , барракуды, корифены и прибрежные виды скумбриевых . Многие из них питаются кормовыми рыбами и в свою очередь служат кормовой базой для крупных пелагических хищников. Почти все рыбы являются хищниками, поэтому, за исключением видов, занимающих верхнее положение в пищевой цепи , разделение на кормовых и хищных рыб до некоторой степени искусственно .
Многие крупные пелагические рыбы являются кочевыми видами, которые совершают длительные морские миграции. Они питаются мелкими и средними пелагическими рыбами. Время от времени, они следуют за добычей, собравшейся в косяки, и многие сами образуют стаи. В омывающих Европу водах существует три популяции скумбрии . Одна популяция мигрирует в Северное море , другая остаётся в Ирландском море , а третья уплывает на юг вдоль западного побережья Шотландии и Ирландии . Скумбрии живут в толще воды близко к поверхности, они постоянно двигаются, перемещаясь между районами нагула и местами нереста. Их промышляют разноглубинными пелагическими тралами, кошельковыми неводами и дрифтерными сетями .
Примерами крупных пелагических рыб являются тунцы, марлины, королевские макрели , акулы и скаты. Тунцы представляют ценный объект для коммерческого промысла. Они бороздят океаны и найти их непросто. Эти рыбы собираются в местах, богатых кормом, вдоль границ течений, у подводных вершин, и в местах апвеллинга. Их ловят кошельковым неводом , пелагическими тралами , ярусами и дрифтерными сетями. Применяют , Сейчас вылавливается около 5 млн тонн тунца, из них 2,3 млн тонн в Тихом океане, уловы постепенно снижаются .
-

-
 Промысел желтопёрого тунца пришёл на смену промысла , численность которого существенно снизилась.
Промысел желтопёрого тунца пришёл на смену промысла , численность которого существенно снизилась. -

-
 Королевская макрель
Королевская макрель
Продуктивность

- Апвеллинг наблюдается как в прибрежной зоне, так и в открыто океане, когда благодаря столкновению глубинных океанических течений, холодные воды, богатые питательными веществами, поднимаются на поверхность. Апвеллинг вызывает цветение планктона, который в свою очередь производит зоопланктон, поддерживающий основные мировые рыбные ресурсы. С прекращением апвеллинга, рыболовство приходит в упадок .
В 1960-х годах промысел перуанского анчоуса был крупнейшим в мире. Численность анчоуса сильно снизилась во время Эль-Ниньо 1972 года, когда тёплая вода прошла над холодным течением Гумбольдта , как часть 50-летнего цикла, понизив глубину термоклина. Апвеллинг остановился, производство фитопланктона упало, вслед за этим снизилась численность популяции анчоуса, и погибли миллионы птиц, которым он служил кормовой базой . В середине 80-х годов апвеллинг возобновился, и уловы перуанского анчоуса вернулись к прежнему уровню.
У берегов Японии столкновение холодного Курильского течения и тёплого течения Куросио вызывает богатый питательными веществами апвеллинг. Циклические изменения этих течений привели к снижению численности популяции перуанской сардины . Уловы упали с 5 миллионов тонн в 1988 году до 280 тысяч тонн в 1998 году. В результате в регион перестали приходить на нагул тихоокеанские голубые тунцы .
Океанические течения могут влиять на распределение рыбных ресурсов, как концентрируя, так и рассеивая их. Смежные течения имеют чёткие границы. Иногда они даже видимы, хотя в большинстве случаев их легко определить по изменению солёности, температуры и мутности воды .
Например, в азиатской части северной Пацифики длиннопёрые тунцы встречаются между двумя системами течений. Северная граница их ареала очерчена холодным Северо-Тихоокеанским течением , а южная — Североэкваториальным . На их распределение в очерченной области оказывает влияние течение Куросио, подверженное сезонным колебаниям .
Острова и банки могут взаимодействовать с течениями и апвеллингом, создавая области с высокой продуктивностью. Большие завихрения создают от островов нисходящие или подветренные течения, в которых концентрируется планктон . Банки и рифы могут стать причиной резкого подъёма глубоких течений .
Далеко мигрирующие виды рыб
Эпипелагические рыбы в целом часто перемещаются на большие расстояния между местами нагула и нереста, или в реагируя на изменения окружающей среды. Крупные океанические хищники, такие как тунцы и лососи, способны преодолеть тысячи километров, пересекая океаны .
В 2001 году было произведено исследование миграций обыкновенного тунца от побережья Северной Каролины с помощью специальных всплывающих маячков. После прикрепления к телу маячок отслеживал перемещения рыбы в течение года, после чего отцеплялся, всплывал на поверхность воды и передавал данные на спутник . Исследование показало, что существует 4 модели миграций. Одна группа тунцов целый год держалась в рамках западной Атлантики. Вторая большую часть времени также провела в западной части Атлантического океана, но на нерест уплыла в Мексиканский залив . Третья группа пересекла Атлантику и возвратилась обратно. Тунцы, принадлежащие к четвёртой группе, также переплыли через Атлантический океан и зашли на нерест в Средиземное море . Исследование показало, что несмотря на разные районы нереста, в северной Атлантике существует единая популяция обыкновенного тунца, а группы перемешиваются между собой .
Мигрирующие виды — официальный термин, принятый в статье 64 Конвенции ООН по морскому праву . Это тунцы и близкородственные виды (длиннопёрый, обыкновенный, большеглазый , полосатый , желтопёрый , чернопёрый , , тихоокеанский голубой, малые тунцы и макрелетунцы , морские лещи , марлины , парусники , меч-рыба , карликовая сайра , океанические акулы, дельфины и другие китообразные . По существу к мигрирующим видам относятся все крупные пелагические рыбы, за исключением не представляющих коммерческой ценности, например, луны-рыбы . Эти виды занимают высокое положение в трофической цепи и совершают длительные миграции к местам нереста и нагула, зачастую следуя за кормовым видами, а также имеют обширное географическое распространение. Таким образом эти виды встречаются как внутри исключительной экономической зоны (370 км от берега), так и в открытом море за её пределами. Это пелагические виды, которые не держатся у дна, хотя определённую часть жизненного цикла они могут проводить в прибрежных водах .
Примечания
- ↑ Lal, Brij V.; Fortune, Kate. . — University of Hawaii Press, 2000. — P. . — ISBN 978-0-8248-2265-1 .
- . www.seapeace.ru. Дата обращения: 25 декабря 2017. 17 мая 2017 года.
- ↑ , pp. 585.
- McLintock, Alexander Hare, Lawrence James Paul, B.SC.,. (англ.) . Fisheries Division, Marine Department, Wellington., New Zealand Ministry for Culture and Heritage Te Manatu Taonga. Дата обращения: 31 января 2016. 24 октября 2007 года.
- Walrond, Carl. (англ.) . Te Ara Encyclopedia of New Zealand . New Zealand Ministry for Culture and Heritage Te Manatu Taonga. Дата обращения: 31 января 2016. 22 октября 2014 года.
- . Технология рыбы и рыбных продуктов. Fish-industry.ru. Дата обращения: 31 января 2016. 10 октября 2016 года.
- (англ.) в базе данных FishBase .
- А. А. Иванов. Физиология рыб / Под ред. С. Н. Шестах. — М. : Мир, 2003. — 284 с. — (Учебники и учебные пособия для студентов высших учебных заведений). — 5000 экз. — ISBN 5-03-003564-8 .
- Наумов Н. П., Карташев Н. Н. Часть 1.Низшие хордовые, бесчелюстные, рыбы, земноводные // Зоология позвоночных. — М. : Высшая школа, 1979. — С. 117—118.
- ↑ Экологический энциклопедический словарь / Под редакцией И.И. Дедю. — Кишинёв, 1989.
- Словарь морских терминов от 31 января 2016 на Wayback Machine
- ↑ , pp. 571.
- Котт К. Приспособительная окраска животных. — Москва, 1950.
- Микулин А. Е., Черняев Ж. А. . Стенограмма эфира программы Гордона. Диалоги (август 2003 г.) . gordon0030.narod.ru. Дата обращения: 31 января 2016. 26 апреля 2016 года.
- Сабунаев В. Б. // Занимательная ихтиология. — 1967. 9 августа 2016 года.
- Herring, Peter. (англ.) . — Oxford University Press, 2002. — P. —195. — ISBN 978-0-19-854956-7 .
- ↑ , pp. 572.
- Blackburn. Oceanography and the ecology of tunas // Oceanography and Marine Biology: An Annual Review. — 1965. — Т. 3 . — С. 299—322 .
- Hunter, JR; Mitchell CT. Association of fishes with flotsam in the offshore waters of Central America // US Fishery Bulletin. — 1966. — Т. 66 . — С. 13—29 .
- Kingsford M. J. Biotic and abiotic structure in the pelagic environment: Importance to small fishes (англ.) // Bulletin of Marine Science. — , 1993. — Vol. 53 , no. (2) . — P. 393—415 .
- Dooley J. K. Fishes associated with the pelagic sargassum complex, with a discussion of the sargassum community // Contributions in Marine Science. — 1972. — № 16 . — С. 1—32 .
- , pp. 576.
- ↑ , pp. 574—575.
- Erwan Josse, Laurent Dagorn, Arnaud Bertrand. // Aquatic Living Resources. — 2000-07-01. — Vol. 4, № 13 . — P. 183—192. — doi : .
- John G. Frazier, Harry L. Fierstine, Sallie C. Beavers, Federico Achaval, Hiroyuki Suganuma. (англ.) // Environmental Biology of Fishes. — 1994-01-01. — Vol. 39 , iss. 1 . — P. 85—96 . — ISSN . — doi : . 10 июня 2018 года.
- ↑ , pp. 591.
- ↑ R. L. Haedrich. (англ.) // . — Wiley-Blackwell , 1996-12-01. — Vol. 49 . — P. 40—53 . — ISSN . — doi : . 13 декабря 2014 года.
- , pp. 586.
- ↑ Ryan P. (англ.) . Te Ara Encyclopedia of New Zealand . New Zealand Ministry for Culture and Heritage Te Manatu Taonga. Дата обращения: 5 февраля 2016. 13 января 2016 года.
- E. L. Douglas, W. A. Friedl, G. V. Pickwell. (англ.) // Science. — 1976-03-05. — Vol. 191 , iss. 4230 . — P. 957—959 . — ISSN . — doi : . 10 апреля 2016 года.
- , pp. 590.
- W. R. A Muntz. // Journal of the Marine Biological Association of the United Kingdom. — 1976-11-01. — Vol. 04, № 56 . — P. 963—976. — ISSN . — doi : .
- (англ.) в базе данных FishBase .
- от 9 октября 2015 на Wayback Machine ScienceDaily , 24 February 2009.
- ↑ Kenaley, C.P. Revision of the Stoplight Loosejaw Genus Malacosteus (Teleostei: Stomiidae: Malacosteinae), with Description of a New Species from the Temperate Southern Hemisphere and Indian Ocean (англ.) // : journal. — 2007. — Vol. 2007 , no. 4 . — P. 886—900 . — doi : .
- Sutton, T.T. Trophic ecology of the deep-sea fish Malacosteus niger (Pisces: Stomiidae ): An enigmatic feeding ecology to facilitate a unique visual system? (англ.) // Deep Sea Research Part I: Oceanographic Research Papers : journal. — 2005. — Vol. 52 , no. 11 . — P. 2065—2076 . — doi : .
- Hans-Joachim Wagner, Ron H. Douglas, Tamara M. Frank, Nicholas W. Roberts, Julian C. Partridge. (англ.) // Current Biology . — Cell Press , 2009-01-27. — Vol. 2 , no. 19 . — P. 108—114 . — ISSN . — doi : .
- . www.americanscientist.org. Дата обращения: 6 февраля 2016. Архивировано из 6 февраля 2016 года.
- Hulley, P. Alexander Paxton, J.R. & Eschmeyer, W.N., ed. . — San Diego: Academic Press, 1998. — P. 127—128. — ISBN 0-12-547665-5 .
- (pdf). 2006 SCOR General Meeting . SCOR. Дата обращения: 6 февраля 2016. 6 февраля 2016 года.
- Dr P.A. (Butch) Hulley. . Natural History Research . Iziko museums of South Africa. Дата обращения: 6 февраля 2016. Архивировано из 16 января 2017 года.
- J.D. Knight. . www.seasky.org. Дата обращения: 6 февраля 2016. 1 февраля 2016 года.
- Kurt M. Schaefer, Daniel W. Fuller. (англ.) // Marine Biology. — 2010-08-10. — Vol. 12 , no. 157 . — P. 2625—2642 . — ISSN . — doi : . 6 сентября 2017 года.
- , pp. 336.
- (англ.) в базе данных FishBase .
- (англ.) в базе данных FishBase .
- ↑ New Zealand Ministry for Culture and Heritage Te Manatu Taonga. (англ.) . www.teara.govt.nz. Дата обращения: 6 февраля 2016. 6 января 2016 года.
- Батипелагиаль // Геологический словарь. Т. 1. М.: Недра, 1978.
- ↑ , pp. 594.
- Ryan P. (англ.) . www.teara.govt.nz. Дата обращения: 5 февраля 2016. 13 января 2016 года.
- (англ.) в базе данных FishBase .
- ↑ , pp. 587.
- Randolph E. Schmid. . Associated Press. Дата обращения: 6 февраля 2016. 4 марта 2016 года.
- Marshall. Progenetic tendencies in deep-sea fishes. — in Potts G. W. and Wootton R. J. (eds.) Fish reproduction: strategies and tactics Fisheries Society of the British Isles. — 1984. — P. 91—101.
- Horn, M H. (англ.) // Breviora : journal. — 1970. — Vol. 359 . — P. 1—9 . 1 мая 2015 года. .
- George Y. Jumper, Ronald C. Baird. (англ.) // The American Naturalist . — University of Chicago Press , 1991. — Vol. 6 , no. 138 . — P. 1431—1458 . — ISSN . — doi : . 1 августа 2020 года.
- Theodore W. Pietsch. (англ.) // Nature. — 1975-07-03. — Vol. 256 , iss. 5512 . — P. 38—40 . — doi : . 28 августа 2008 года.
- Jordan, D.S. . — H. Holt and Company, 1905.
- (англ.) в базе данных FishBase .
- (англ.) в базе данных FishBase .
- (англ.) в базе данных FishBase .
- . Центр системы мониторинга рыболовства и связи. Дата обращения: 7 февраля 2016. Архивировано из 15 марта 2016 года.
- ↑ Walrond C. Carl. (англ.) . Te Ara Encyclopedia of New Zealand . New Zealand Ministry for Culture and Heritage Te Manatu Taonga. Дата обращения: 7 февраля 2016. 3 февраля 2016 года.
- ↑ , pp. 588.
- J. Mauchline, J. D. M. Gordon. // Marine Ecology Progress Series. — № 27 . — P. 227—238. — doi : . 3 марта 2016 года.
- J. A. Koslow. (англ.) // . — Wiley-Blackwell , 1996-12-01. — Vol. 49 . — P. 54—74 . — ISSN . — doi : . 13 декабря 2014 года.
- (англ.) в базе данных FishBase .
- ↑ M. L. Fine, M. H. Horn, B. Cox. (англ.) // Proceedings of the Royal Society of London B: Biological Sciences. — 1987-03-23. — Vol. 230 , iss. 1259 . — P. 257—265 . — ISSN . — doi : . 7 февраля 2016 года.
- . Регулятивная система . Commission for the Conservation of Antarctic Marine Living Resources. Дата обращения: 7 февраля 2016. 21 января 2016 года.
- . www.inokean.ru. Дата обращения: 7 февраля 2016. 26 октября 2015 года.
- Sulak KJ. // Ichthyological Research. — Т. 32 , № (4) . — С. 443—446 . 16 февраля 2016 года.
- ↑ Nielsen J. G. // Galathea Report. — 1977. — Т. 14 . — С. 41—48 . 15 февраля 2016 года.
- (англ.) в базе данных FishBase .
- (англ.) в базе данных FishBase .
- Checkley D., Alheit J. and Oozeki Y. Climate Change and Small Pelagic Fish. — Cambridge University Press, 2009. — ISBN 0-521-88482-9 .
- . www.iznorvegii.ru. Дата обращения: 18 августа 2016. 31 июля 2016 года.
- . www.wwf.ru. Дата обращения: 18 августа 2016. Архивировано из 1 сентября 2016 года.
- . www.wwf.ru. Дата обращения: 18 августа 2016. Архивировано из 30 сентября 2016 года.
- . www.fao.org. Дата обращения: 11 февраля 2016. 21 декабря 2018 года.
- . Institute of Marine Research. Дата обращения: 11 февраля 2016. 11 августа 2017 года.
- . www.wwf.ru. Дата обращения: 11 февраля 2016. Архивировано из 15 февраля 2016 года.
- . www.wwf.ru. Дата обращения: 11 февраля 2016. Архивировано из 15 февраля 2016 года.
- Francisco P. Chavez, John Ryan, Salvador E. Lluch-Cota, Miguel Ñiquen C. (англ.) // Science. — 2003-01-10. — Vol. 299 , iss. 5604 . — P. 217—221 . — ISSN . — doi : . 3 июля 2016 года.
- Jeffrey J. Polovina. (англ.) // Fisheries Oceanography. — 1996-06-01. — Vol. 5 , iss. 2 . — P. 114—119 . — ISSN . — doi : . 13 декабря 2014 года.
- . www.fao.org. Дата обращения: 18 февраля 2016. 24 сентября 2015 года.
- Hiroshi Nakamura. Tuna distribution and migration. — Fishing News. — Nature, 1969.
- Blackburn M. // Oceanography and Marine Biology Annual Revue. — 1965. — № 3 . — С. 299—322 . 3 марта 2016 года.
- , pp. 578.
- Barbara A. Block, Heidi Dewar, Susanna B. Blackwell, Thomas D. Williams, Eric D. Prince. (англ.) // Science. — 2001-08-17. — Vol. 293 , iss. 5533 . — P. 1310—1314 . — ISSN . — doi : . 10 марта 2016 года.
- . www.un.org. Дата обращения: 18 февраля 2016. 2 сентября 2017 года.
- NOAA Fisheries West Coast Region. (англ.) . www.westcoast.fisheries.noaa.gov. Дата обращения: 18 февраля 2016. 4 февраля 2016 года.
Литература
- Moyle, P. B. and Cech, J. J. . — Benjamin Cummings, 2004. — ISBN 978-0-13-100847-2 .
Ссылки
- 2020-01-14
- 1












