Факторы транскрипции
- 1 year ago
- 0
- 0
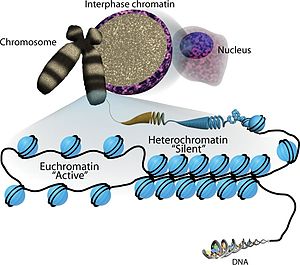
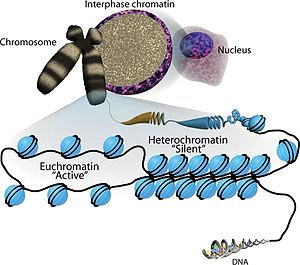
Эухромати́н , также активный или «открытый хромати́н» — участки хроматина , которые представляет собой неплотную и легко упакованную форму ( ДНК , РНК и белки), обогащённую генами и часто (но не всегда) находящуюся в состоянии активной транскрипции . Эухроматин отличается от гетерохроматина , который плотно упакован и менее доступен для транскрипции. Около 92 % генома человека является эухроматичным .
У эукариот эухроматин представляет собой наиболее активную часть генома в клеточном ядре . У прокариот эухроматин является единственной формой хроматина; это указывает на то, что гетерохроматиновая структура развилась позже вместе с ядром, возможно, как механизм, позволяющий справиться с увеличением размера генома.
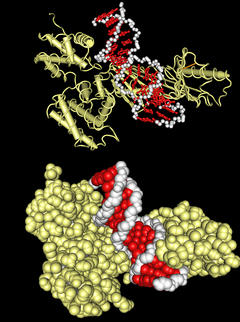
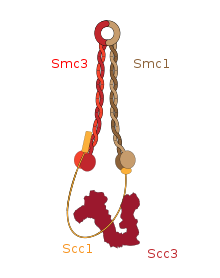
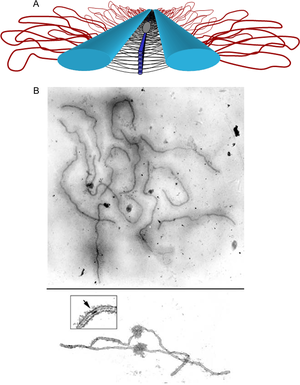
Эухроматин состоит из повторяющихся нуклеопротеидных субъединиц, известных как нуклеосомы , напоминающие развёрнутый набор бусин на нитке, диаметром около 11 нм . В основе этих нуклеосом находится набор из четырёх пар белков- гистонов : H3, H4, H2A и H2B . Каждый оснóвный гистоновый белок имеет "хвостовую" структуру, которая может изменяться несколькими способами; считается, что эти вариации действуют как "главные переключатели управления" через различные состояния метилирования и ацетилирования, которые определяют общее расположение хроматина . Вокруг гистоновых октамеров намотано около 147 пар оснований ДНК , что составляет чуть меньше 2 витков спирали . Нуклеосомы вдоль нити связаны между собой гистоном H1 и коротким участком открытой линкерной ДНК, составляющим около 0-80 пар оснований. Ключевое различие между структурой эухроматина и гетерохроматина заключается в том, что нуклеосомы в эухроматине расположены гораздо шире, что обеспечивает более лёгкий доступ различных белковых комплексов ( транскрипционных факторов ) к нити ДНК и тем самым повышает транскрипцию генов .
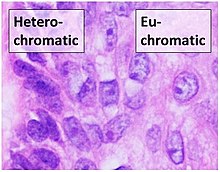
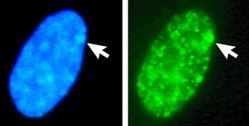
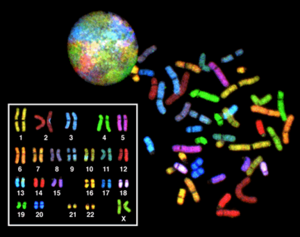
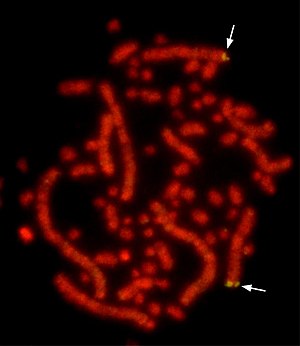
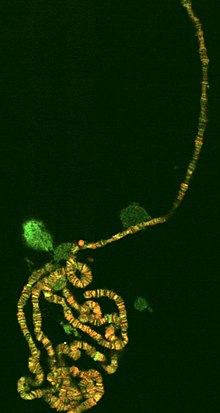
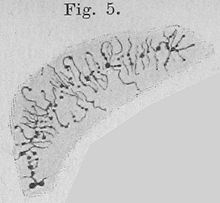
Эухроматин при большом увеличении напоминает набор бусинок на нитке . С более дальнего расстояния он может напоминать клубок запутанных ниток, как, например, на некоторых изображениях, полученных с помощью электронного микроскопа . Как при оптической, так и при электронной микроскопии эухроматин выглядит светлее, чем гетерохроматин, который также присутствует в ядре и выглядит тёмным , из-за его менее компактной структуры . При визуализации хромосом , например, на кариограмме, для окрашивания используется цитогенетическое окрашивание. Цитогенетическое окрашивание позволяет нам увидеть, какие части хромосомы состоят из эухроматина или гетерохроматина, чтобы дифференцировать хромосомные участки, нарушения или перестройки. . Одним из таких примеров является G-бэндинг, иначе известное как окрашивание по Гимзе, при котором эухроматин кажется светлее, чем гетерохроматин .
| Окрашивание по Гимзе (G-) бэндинг | Реверсивное или обратное окрашивание (R-) бэндинг | Конститутивный гетерохроматин (C-) бэндинг | Акрихиновое окрашивание (Q-) бэндинг | Теломерное окрашивание (T-) бэндинг | |
|---|---|---|---|---|---|
| Эухроматин | Светлее | Темнее | Светлее | Тусклее | Светлее |
| Гетерохроматин | Темнее | Светлее | Темнее | Ярче (флуоресцентное свечение) | Темнее (слабо отчётливо) |

Эухроматин участвует в активной транскрипции ДНК в молекулы мРНК . Развёрнутая структура эухроматина позволяет белкам генной регуляции и комплексам РНК-полимеразы связываться с последовательностью ДНК, что может инициировать процесс транскрипции . Хотя не весь эухроматин обязательно транскрибируемый, поскольку он разделён на транскрипционно активные и неактивные домены , эухроматин всё же обычно ассоциируется с активной транскрипцией генов. Поэтому существует прямая связь между тем, насколько активно продуктивна клетка, и количеством эухроматина, которое можно обнаружить в её ядре .
Считается, что клетка использует трансформацию (преобразование) эухроматина в гетерохроматин как метод контроля экспрессии и репликации генов, поскольку эти процессы по-разному протекают в достаточно плотном хроматине. Это явление известно как " гипотеза доступности " . Одним из примеров конститутивного эухроматина, который "всегда включён", являются гены домашнего хозяйства , которые кодируют белки, необходимые для основных функций выживания и функционирования клетки .
Эпигенетика подразумевает изменения в фенотипе , которые могут быть унаследованы без изменения последовательности ДНК. Это может происходить в результате многих типов взаимодействия с окружающей средой . Что касается эухроматина, посттрансляционные модификации гистонов могут изменять структуру хроматина, что приводит к изменению экспрессии генов без изменения молекул ДНК . Кроме того, было показано, что потеря гетерохроматина и увеличение эухроматина коррелируют с ускорением процесса старения, особенно при заболеваниях, похожих на прогерию (преждевременное старение) . Исследования показали наличие эпигенетических маркеров на гистонах для ряда других заболеваний .
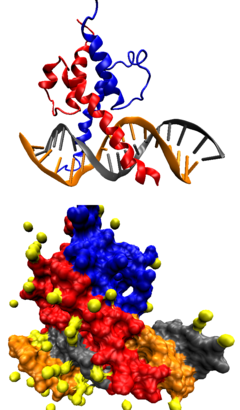
Регуляция эухроматина происходит в первую очередь посттрансляционными модификациями гистонов нуклеосом, осуществляемыми многими гистон-модифицирующими ферментами. Эти модификации происходят на N-концевых хвостах гистонов, выступающих из структуры нуклеосом, и, как полагают, рекрутируют ферменты для поддержания хроматина либо в открытой форме, как эухроматин, либо в закрытой форме, как гетерохроматин . Ацетилирование гистонов, например, обычно связано со структурой эухроматина, тогда как метилирование гистонов способствует ремоделированию гетерохроматина . Ацетилирование делает группу гистонов более отрицательно заряженной, что, в свою очередь, нарушает её взаимодействие с нитью ДНК, по сути "открывая" нить, облегчая к ней доступ . Ацетилирование может происходить на нескольких лизиновых остатках N-концевого хвоста гистона и на разных гистонах одной нуклеосомы, что, как считается, ещё больше увеличивает доступность ДНК для факторов транскрипции .
Фосфорилирование гистонов — ещё один способ регуляции эухроматина . Как правило, оно происходит на N-концевых хвостах гистонов, однако некоторые участки присутствуют и в кóровых гистонах . Фосфорилирование контролируется киназами и фосфатазами , которые добавляют и удаляют фосфатные группы соответственно. Оно может происходить на остатках серина , треонина или тирозина , присутствующих в эухроматине . Поскольку фосфатные группы, добавленные к молекулам гистонов, несут отрицательный заряд, это способствует образованию более расслабленной "открытой" формы, подобно ацетилированию . Что касается функциональности, фосфорилирование гистонов участвует в экспрессии генов , репарации повреждений ДНК и ремоделировании хроматина .
Другой метод регуляции, который связан с присоединением отрицательного заряда, благоприятствующего "открытой" форме — это АДФ-рибозилирование . В данном процессе происходит присоединение одной или несколько единиц АДФ-рибозы к гистону, также этот процесс участвует в ответной реакции на повреждение ДНК .
|
|
|
|---|---|
| Словари и энциклопедии |
|
| Основное |

|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
|
||||||||||
|
Перестройки и
нарушения |
|||||||||||
|
Хромосомное
определение пола |
|||||||||||
| Методы | |||||||||||