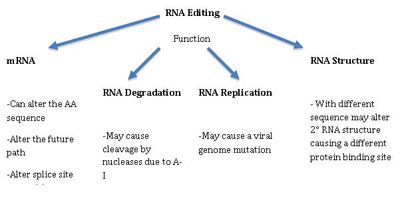
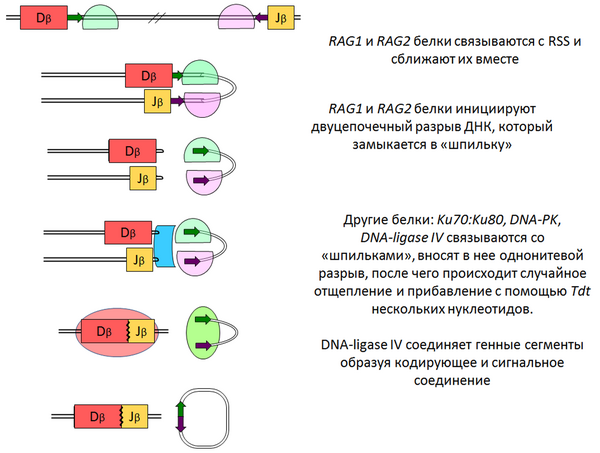
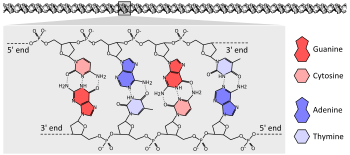
Мутация
- 1 year ago
- 0
- 0

Фреймшифт-мутация, мутация сдвига рамки считывания — тип мутации в последовательности ДНК , для которого характерна вставка или делеция нуклеотидов , в количестве не кратном трём. В результате происходит сдвиг рамки считывания при транскрипции мРНК . Мутации сдвига рамки считывания делятся на мишенные, немишенные, мишенные задерживающиеся и немишенные задерживающиеся мутации сдвига рамки считывания.
Следует различать мутацию сдвига рамки считывания, в которой происходит вставка или делеция нуклеотида, от однонуклеотидного полиморфизма , в котором происходит замена одного нуклеотида на другой.
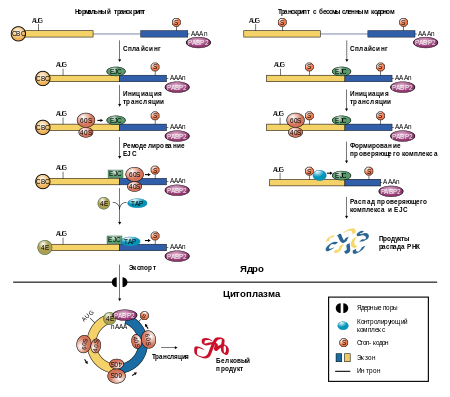
В связи с триплетным характером генетического кода , вставка или делеция числа нуклеотидов, не кратных трём, приводит к сильному искажению информации в транскрибируемой мРНК. Также в результате может появиться стоп-кодон , что приводит к преждевременной терминации синтеза белка.
Также может возникнуть и обратная ситуация, когда изменённый стоп-кодон начинает кодировать какую-либо аминокислоту. Это приводит к аномальному удлинению полипептидной цепи. Когда делеция и вставка кодонов происходят друг за другом последовательно в одной и той же точке цепи кодонов в ДНК, это может привести к синтезу белка нужной длины, но с другой аминокислотой в измененном фрагменте (SNP-мутация — однонуклеотидный полиморфизм ).
Мишенная мутация сдвига рамки считывания — это мутация сдвига рамки считывания, которая появляется напротив повреждений ДНК, способных останавливать синтез ДНК. Например, напротив циклобутановых пиримидиновых димеров — основной причине ультрафиолетового мутагенеза. Термин происходит от слова «мишень». Часть мишенных мутаций сдвига рамки, например, инсерции и делеции в один нуклеотид, можно отнести к точечным мутациям. Они делятся на мишенные делеции, мишенные инсерции, мишенные сложные делеции и мишенные сложные инсерции и, соответственно, на мишенные задерживающиеся делеции, мишенные задерживающиеся инсерции, мишенные задерживающиеся сложные делеции и мишенные задерживающиеся сложные инсерции .
В настоящее время наиболее обоснованной моделью, объясняющей механизм образования мутаций сдвига рамки, считается модель Стрейзингера , которая предполагает, что причина образования мутаций сдвига рамки лежит в появлении брешей и проскальзывании нити ДНК во время синтеза . Было показано, что образование делеций связано с появлением петель или выпуклостей в молекуле ДНК .
В рамках полимеразно-таутомерной модели ультрафиолетового мутагенеза были развиты модели механизмов образования мишенных инсерций , мишенных делеций и мишенных сложных инсерций , вызванных цис-син-циклобутановыми тиминовыми димерами. Структурный анализ встраивания канонических оснований ДНК напротив цис-син-циклобутановых тиминовых димеров, содержащих тимин в одной определенной редкой таутомерной форме, показал, что напротив них невозможно встроить ни одно каноническое основание так, чтобы образовались водородные связи между основаниями в этой редкой таутомерной форме, и каноническими основаниями ДНК. Напротив цис-син-циклобутановых тиминовых димеров, содержащих молекулы тимина в этой редкой таутомерной форме, могут появиться бреши в один нуклеотид. Проскальзывание нити ДНК и образование петель может привести к образовании делеций или инсерций.