Interested Article - Классический путь активации системы комплемента
- 2021-08-06
- 1

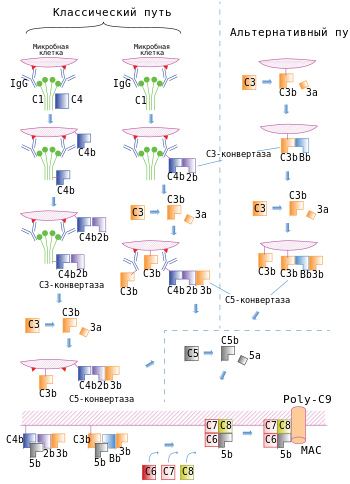
Класси́ческий пу́ть актива́ции систе́мы комплеме́нта — один из трёх путей активации системы комплемента , наряду с альтернативным путём и лектиновым путём . Классический путь запускают комплексы антиген - антитело ( ), причём антитела должны принадлежать к классу IgG или IgM . После активации происходит сборка белкового (C4b2b), который разрезает белок комплемента . Один из фрагментов C3, , связывается с C3-конвертазой, в результате чего образуется (C4b2b3b). C5-конвертаза разрезает компонент комплемента 5, после чего образовавшиеся белковые фрагменты привлекают фагоциты в очаг инфекции и способствуют поглощению клеток патогенов . C5-конвертаза также запускает финальные стадии каскада комплемента, которые завершаются формированием мембраноатакующего комплекса. Он формирует поры в мембране клетки микроорганизма , вызывая её лизис и гибель. Помимо микробных клеток, классический путь комплемента могут запускать апоптотические тельца и некротизированные клетки .
Классический путь активации комплемента был открыт в 1888 году Жюлем Борде и стал первым описанным механизмом активации системы комплемента .
Механизм
Инициация
Классический путь инициируется при связывании с доменом C H 2 молекулы иммуноглобулина G (IgG) или доменом C H 3 молекулы иммуноглобулина M (IgM), которые уже связались с антигеном. Среди антител IgG классический путь наиболее эффективно активируют IgG3 и IgG1 (у человека). Белок C1 состоит из субъединиц , и , причём C1q связывается с антителом, а C1r и C1s являются протеазами . C1q представляет собой гексамер , который специфически связывается с участками Fc тяжёлых цепей типа μ и некоторыми тяжёлыми цепями типа γ. Классический путь комплемента не может быть активирован свободными антителами, но только антителами, связанными с соответствующим антигеном, причём для активации необходимо, чтобы C1 связался с двумя и более участками Fc. Поскольку каждая молекула IgG имеет только один участок Fc, для связывания с C1 необходимо, чтобы две или более молекулы IgG находились рядом. Хотя свободные IgM в плазме крови являются , путь комплемента не может быть активирован при связывании лишь с одной молекулой IgM, поскольку участки Fc каждого мономера располагаются так, что они не могут быть связаны одной молекулой C1. Одна молекула IgM, будучи пентамером, может связать две молекулы C1, поэтому IgM активирует комплемент эффективнее, чем IgG. C1r и C1s являются сериновыми протеазами и формируют тетрамер , в котором C1r и C1s имеются в числе двух молекул .
Формирование C3-конвертазы
Когда C1q связывается с IgG или IgM, связанный C1r активируется и вносит разрез в C1s, активируя его. Активированный C1s разрезает следующий белок каскада, , с образованием C4b. Как и C3b, C4b содержит внутреннюю тиоэфирную связь , которая обеспечивает ковалентное сшивание C4b с комплексом антиген-антитело на поверхности микробной клетки или непосредственно с поверхностью клетки. Следующий участник каскада, , связывается с C4b, ковалентно прикреплённым к поверхности клетки, и разрушается C1s с образованием фрагмента C2b, функции которого неизвестны. При этом C2a остаётся связанным с C4b на поверхности клетки патогена (в отличие от остальных компонентов комплемента, у C2 больший фрагмент называется C2a, а меньший фрагмент C2b, высвобождаемый при расщеплении С2, остаётся несвязанным). Комплекс C4b2a является C3-конвертазой и может связывать C3 и ферментативно расщеплять его. Связывание C3 с C3-конвертазой обеспечивается C4b, а C2a катализирует протеолиз C3. При расщеплении C3 образуются два фрагмента, меньший из которых, C3a, удаляется, а C3b может ковалентно связываться с белками клеточной поверхности или антителами, связанными с клеткой, на поверхности которой был активирован каскад комплемента. C3b также может взаимодействовать с фактором B и образовывать больше C3-конвертаз по альтернативному пути активации комплемента. Одна C3-конвертаза в итоге может давать начало сотням и тысячам молекул C3b на поверхности клетки, где был активирован комплемент. Ранние этапы классического и альтернативного путей комплемента имеют много похожего: C3 в альтернативном пути гомологичен C4 классического пути, а фактор B гомологичен C2. Некоторые молекулы C3b связываются с C3-конвертазой с образованием комплекса C4b2a3b, который является C5-конвертазой. C5-конвертаза расщепляет и запускает поздние стадии каскада комплемента .
При инфекциях , вызванных пневмококками , запускается независимый от антител, но зависимый от C1 вариант классического пути, который активируется при связывании углеводов с лектинами на поверхности клетки. Некоторые макрофаги экспрессируют лектин C-типа , известный как SIGN-R1, который распознаёт полисахариды пневмококков и связывается с C1q. Благодаря этому активируется классический путь комплемента, в результате работы которого клетка пневмококка покрывается C3b .
Формирование C5-конвертазы и мембраноатакующего комплекса
C5-конвертазы, образовавшиеся в ходе классического, альтернативного или лектинового пути, запускают последующие этапы каскада комплемента, кульминацией которых является образование мембраноатакующего комплекса. C5-конвертаза расщепляет C5 на высвобождаемый меньший фрагмент и больший фрагмент C5b, который остаётся связанным с белками комплемента на поверхности микробной клетки. Последующие участники каскада комплемента — , , и — являются структурно близкими белками, лишёнными ферментативной активности. C5b временно сохраняет конформацию , в которой он может связать C6 и C7 с образованием комплекса C5b,6,7. C7 обладает гидрофобными свойствами и вставляется в липидный бислой клеточной мембраны, где становится высокоаффинным рецептором C8. Белок C8 имеет структуру, и одна из его субъединиц связывается с комплексом C5b,6,7, при этом формируя ковалентную связь со второй субъединицей; третья субъединица интегрирует в клеточную мембрану. Получившийся в результате комплекс C5b,6,7,8 (C5b-8) обладает невысокой способностью к лизису клетки, и формирование полностью функционального мембраноатакующего комплекса завершается при связывании с C5b,6,7,8 компонента C9. C9 полимеризуется в местах взаимодействия с комплексом C5b,6,7,8 и формирует поры в мембране. Через поры, имеющие диаметр около 100 ангстрем , свободно перемещаются вода и ионы . Вход воды в клетку из-за осмоса приводит к её набуханию и разрушению. Поры, сформированные C9, похожи на поры, образованные белком перфорином , который входит в состав гранул цитотоксических T-лимфоцитов и натуральных киллеров , кроме того, C9 структурно гомологичен перфорину .
Клиническое значение
Недостаточность по белку C1q может стать причиной развития системной красной волчанки . Помимо других функций, C1q запускает удаление из тканей и сосудов апоптотических телец и иммунных комплексов . При его недостаточной активности иммунные комплексы и апоптотические тельца накапливаются, вызывая воспаление и аутоиммунные процессы, при которых образуются аутоантитела . Изучается возможность использования аутоантител к C1q в качестве системной красной волчанки .
Чрезмерная активность классического пути комплемента при недостаточной работе может приводить к эпизодической ангиоэдеме . Недостаточность по C1-ингибитору может быть наследственной или приобретённой . В норме C1-ингибитор инактивирует C1r и C1s, мешая запуску классического пути активации комплемента. Кроме того, C1-ингибитор контролирует проницаемость сосудов. Концентрация C1-ингибитора, составляющая 50 % и менее от нормальной, приводит к повышению проницаемости сосудов, из-за чего развивается ангиоэдема . В 2008 году для предотвращения приступов наследственной ангиоэдемы был одобрен препарат цинриз ( англ. Cinryze ), представляющий собой производное C1-ингибитора из плазмы крови человека .
Изучается возможность уничтожения вирионов ВИЧ с помощью классического пути активации комплемента . Показана эффективность методов иммунотерапии рака , использующих активацию классического пути . Классический путь комплемента особенно важен для уничтожения клеток метициллинрезистентных штаммов золотистого стафилококка , так как с ними связываются некоторые варианты IgM .
Примечания
- ↑ Noris M. , Remuzzi G. (англ.) // Seminars In Nephrology. — 2013. — November ( vol. 33 , no. 6 ). — P. 479—492 . — doi : . — .
- Nesargikar P. N. , Spiller B. , Chavez R. (англ.) // European Journal Of Microbiology & Immunology. — 2012. — June ( vol. 2 , no. 2 ). — P. 103—111 . — doi : . — .
- ↑ Thielens N. M. , Tedesco F. , Bohlson S. S. , Gaboriaud C. , Tenner A. J. (англ.) // Molecular Immunology. — 2017. — September ( vol. 89 ). — P. 73—83 . — doi : . — .
- ↑ Vignesh P. , Rawat A. , Sharma M. , Singh S. (англ.) // Clinica Chimica Acta; International Journal Of Clinical Chemistry. — 2017. — February ( vol. 465 ). — P. 123—130 . — doi : . — .
- , с. 167.
- ↑ , p. 276—278.
- , p. 278.
- , p. 279—280.
- Stegert M. , Bock M. , Trendelenburg M. (англ.) // Molecular Immunology. — 2015. — September ( vol. 67 , no. 1 ). — P. 3—11 . — doi : . — .
- Taylor P. R. , Carugati A. , Fadok V. A. , Cook H. T. , Andrews M. , Carroll M. C. , Savill J. S. , Henson P. M. , Botto M. , Walport M. J. (англ.) // The Journal Of Experimental Medicine. — 2000. — 7 August ( vol. 192 , no. 3 ). — P. 359—366 . — doi : . — .
- Chi S. , Yu Y. , Shi J. , Zhang Y. , Yang J. , Yang L. , Liu X. (англ.) // Disease Markers. — 2015. — Vol. 2015 . — P. 450351—450351 . — doi : . — .
- Mahler M. , van Schaarenburg R. A. , Trouw L. A. (англ.) // Frontiers In Immunology. — 2013. — Vol. 4 . — P. 117—117 . — doi : . — .
- ↑ Cugno M. , Zanichelli A. , Foieni F. , Caccia S. , Cicardi M. (англ.) // Trends In Molecular Medicine. — 2009. — February ( vol. 15 , no. 2 ). — P. 69—78 . — doi : . — .
- Lunn M. , Santos C. , Craig T. (англ.) // Journal Of Blood Medicine. — 2010. — Vol. 1 . — P. 163—170 . — doi : . — .
- . Food and Drug Administration (17 июля 2018). Дата обращения: 22 апреля 2020. 22 июля 2017 года.
- Pleguezuelos O. , Stoloff G. A. , Caparrós-Wanderley W. (англ.) // Virology Journal. — 2013. — 4 April ( vol. 10 ). — P. 107—107 . — doi : . — .
- Chen J. , Xu X. M. , Underhill C. B. , Yang S. , Wang L. , Chen Y. , Hong S. , Creswell K. , Zhang L. (англ.) // Cancer Research. — 2005. — 1 June ( vol. 65 , no. 11 ). — P. 4614—4622 . — doi : . — .
- An J. , Li Z. , Dong Y. , Wu J. , Ren J. (англ.) // Biochemical And Biophysical Research Communications. — 2015. — 22 May ( vol. 461 , no. 1 ). — P. 142—147 . — doi : . — .
Литература
- Ярилин А. А. Иммунология. — М. : ГЭОТАР-Медиа, 2010. — 752 с. — ISBN 978-5-9704-1319-7 .
- Abul K. Abbas, Andrew H. Lichtman, Shiv Pillai. Cellular and Molecular Immunology : [ англ. ] . — Philadelphia : Elsevier Saunders, 2015. — ISBN 978-0-323-22275-4 .
| Органы | |||||
|---|---|---|---|---|---|
|
Врождённый
иммунитет |
|
||||
|
Приобретённый
иммунитет |
|
||||
| Медиаторы | |||||
| Иммунный ответ | |||||
|
Иммунологическая
толерантность |
|||||
| Нарушения | |||||
- 2021-08-06
- 1