Interested Article - Морфолиновые олигонуклеотиды

- 2021-10-18
- 1
Морфоли́новые олигонуклеоти́ды , или морфоли́новые олигоме́ры ( англ. morpholino, morpholino oligomer, phosphorodiamidate morpholino oligomer, PMO ) — синтетические олигонуклеотиды , использующиеся в молекулярной биологии для изменения экспрессии генов . Основу молекулярной структуры морфолинового олигонуклеотида составляют метилен морфолиновые кольца и фосфородиамидатные связи . Морфолиновые олигонуклеотиды блокируют доступ других молекул к небольшим (длиной около 25 нуклеотидов ) специфическим последовательностям за счёт комплементарного спаривания с соответствующей РНК . Морфолиновые олигонуклеотиды служат исследовательскими инструментами в обратной генетикe для нокдауна гена .
Нокдаун гена заключается в предотвращении синтеза клеткой соответствующего белка . Морфолиновые олигонуклеотиды также могут влиять на сплайсинг пре- мРНК . Нокдаун гена является мощным средством для изучения функции конкретного гена . Аналогичным образом, вырезание специфического экзона из транскрипта позволяет определить функции, выполняемые соответствующими аминокислотными остатками белка, а иногда приводит к полной инактивации белка. Морфолиновые олигонуклеотиды используются в изучении многих модельных организмов , в том числе мышей , данио-рерио , лягушек и морских ежей .
Разрабатываются пути применения морфолиновых олигонуклеотидов в медицине против патогенных организмов ( бактерий и вирусов ), а также для ослабления проявления генетических заболеваний .
Структура
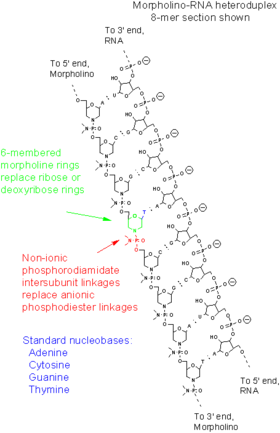
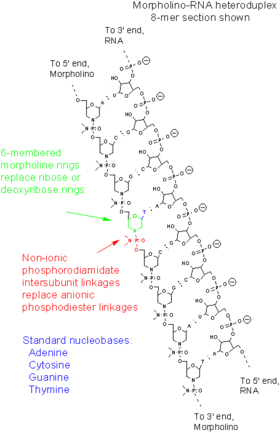
Морфолиновые олигонуклеотиды — это синтетические молекулы, представляющие собой измененные нуклеиновые кислоты . Они, как правило, имеют длину 25 нуклеотидов и комплементарно связываются с соответствующими последовательностями РНК за счёт спаривания оснований . Морфолиновые олигонуклеотиды имеют обычные азотистые основания, которые вместо дезоксирибозы соединены с кольцами морфолина , а в связи отдельных нуклеотидов участвуют не фосфатные , а фосфородиамидатные группы. Замена отрицательно заряженных фосфатных групп незаряженной фосфородиамидатной группой устраняет ионизацию при физиологических значениях рН , так что в живых клетках эти молекулы не заряжены. Вся молекула морфолинового олигонуклеотида состоит из таких изменённых нуклеотидов .
Функции
Механизмы действия морфолиновых олигонуклеотидов
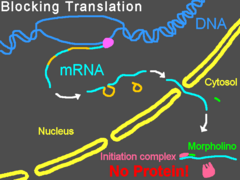
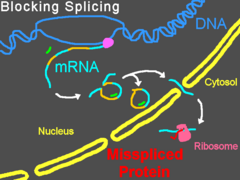
Морфолиновые олигонуклеотиды не вызывают деградации своей РНК-мишени, в отличие от многих антисмысловых молекул (например, малых интерферирующих РНК и ). Вместо этого они выступают в роли «пространственных блокаторов», связываясь с последовательностью-мишенью и физически не давая другим молекулам связаться с этой РНК . Морфолиновые олигонуклеотиды часто используются для изучения роли специфических мРНК в развивающемся зародыше. Эмбриологи инъецируют их в яйца или зародыши данио-рерио , шпорцевой лягушки , морских ежей и рыбы Fundulus heteroclitus или доставляют эти молекулы путём электропорации в зародыши курицы на более поздних стадиях развития ; обработанные таким образом зародыши называют морфантами . При наличии в цитозоле правильных систем доставки морфолиновых олигонуклеотидов могут быть эффективными и в культурах клеток .
Блокирование трансляции
Связываясь с 5'-нетранслируемой областью мРНК, морфолиновые олигонуклеотиды могут мешать рибосоме в комплексе с двигаться от кэпа к старт-кодону . Это предотвращает трансляцию кодирующей области транскрипта-мишени (нокдаун гена). Такой приём очень удобен, когда исследователь хочет определить функцию конкретного белка; по эффектам, оказываемым нокдауном гена на клетку или организм, судят о функции белка. Некоторые морфолиновые олигонуклеотиды блокируют экспрессию столь эффективно, что после деградации белка, синтезированного до введения олигонуклеотидов, этот белок становится неопределимым при помощи вестерн-блоттинга .
Изменение сплайсинга пре-мРНК
Морфолиновые олигонуклеотиды могут вмешиваться в процессинг пре-мРНК следующими способами:
- предотвращая связывание направляющих сплайсинг малых ядерных рибонуклеопротеинов ( ) со своими мишенями на границах интронов в пре-мРНК;
- блокируя остаток аденина , осуществляет нуклеофильную атаку в ходе сплайсинга и образует структуру лассо;
- мешая связыванию регуляторных белков сплайсинга, таких как сайленсеры и .
Предотвращение связывания рибонуклеопротеинов (в сайте-доноре) или / (в или сайте-акцепторе) может привести к модифицированному сплайсингу, при котором из зрелой мРНК исключаются экзоны . Воздействие на другие сайты сплайсинга приводит к включению интронов в зрелую мРНК, в то время как активация скрытых сайтов сплайсинга может приводит и к включениям, и к исключениям . Морфолиновые олигонуклеотиды могут также блокировать мишени snRNP / . Изменения в сплайсинге удобно отслеживать при помощи ПЦР с обратной транскрипцией (RT-PCR) и электрофореза (при электрофорезе продуктов RT-PCR наблюдается смещение полос в геле) .
Другие применения
Морфолиновые олигонуклеотиды использовались для блокировки активности микроРНК , а также их созревания . Морфолиновые олигонуклеотиды, меченные флуоресцеином , в сочетании с антителами , специфичными к флуоресцеину, могут использоваться в качестве проб с микроРНК . Кроме того, они могут блокировать активность рибозимов , а также функционирование snRNP U2 и U12 . Морфолиновые олигонуклеотиды, нацеленные на «оголённые» последовательности мРНК в пределах кодирующей области, могут вызывать при трансляции . Они также могут блокировать редактирование РНК . Эффективность морфолиновых олигонуклеотидов по отношению ко многим мишеням делает их универсальным средством для подавления взаимодействия белков или нуклеиновых кислот с мРНК .
Специфичность, стабильность и не-антисмысловые эффекты
Морфолиновые олигонуклеотиды стали стандартным инструментом для нокдауна генов в эмбриональных системах животных, в которых экспрессируется больше генов, чем во взрослых клетках. После инъекции морфолинового олигонуклеотида в зародыши лягушки или рыбы на стадии одной или нескольких клеток наибольший эффект проявляется к пятому дню , когда большая часть органогенеза и дифференцировки клеток и тканей уже позади; наблюдаемые фенотипы соответствуют нокауту определённого гена. нуклеотиды, не нацеленные на какие-либо последовательности, как правило, не вызывают изменений в фенотипе зародыша, подтверждая, что морфолиновые олигонуклеотиды действуют специфично к последовательностям и, как правило, не имеют не-антисмысловых эффектов. Дозу, необходимую для нокдауна гена, можно сократить, одновременно вводя несколько морфолиновых олигонуклеотидов, нацеленных на одну и ту же мРНК; этот приём позволяет сократить или вовсе исключить зависимые от дозы взаимодействия олигонуклеотидов с РНК, не являющимися непосредственными мишенями .
В экспериментах по «спасению» мРНК у зародышей часто удавалось вернуть фенотип дикого типа . В ходе этих экспериментов морфолиновый олигонуклеотид вводится в клетку вместе с мРНК, кодирующей тот белок, на нокдаун которого этот олигонуклеотид и направлен. Однако вводимая мРНК имеет изменённую 5'-нетранслируемую область , а потому не имеет последовательности-мишени для морфолинового олигонуклеотида, но её кодирующая область сохранена и с неё синтезируется интересующий белок. Трансляция с мРНК-«спасателя» восстанавливает образование белка, остановленное морфолиновым олигонуклеотидом. Спасательная мРНК не вызывает фенотипических отклонений, так как не действует на экспрессию генов, не являющихся мишенью введенного олигонуклеотида, поэтому возврат к фенотипу дикого типа служит ещё одним доказательством специфичности морфолиновых олигонуклеотидов по отношению к последовательности .
Из-за совершенно неестественного химического строения морфолиновые олигонуклеотиды не распознаются клеточными белками. Нуклеазы их не разрушают , поэтому они не разрушаются ни в клетках, ни в плазме крови . Морфолиновые олигонуклеотиды не активируют Toll-подобные рецепторы , а потому не запускают врождённый иммунный ответ , выражающийся, в частности, в образовании интерферона или в NF-κB -опосредованном воспалении .
До 18 % морфолиновых олигонуклеотидов вызывают не связанные с исходными мишенями фенотипы, в частности, смерть клеток центральной нервной системы и сомитов у зародышей данио-рерио . Было показано, что большая часть этих эффектов связана с активацией p53 -опосредованного апоптоза ; их можно подавить, одновременно вводя экспериментальный морфолиновый олигонуклеотид и его анти-p53-аналог. Более того, вызванную морфолиновым олигонуклеотидом активацию p53-опосредованного апоптоза удалось повторить с использованием других антисмысловых структур, что говорит в пользу того, что p53-опосредованный апоптоз может быть вызван утратой белка-мишени и не зависит от типа олигонуклеотида, используемого для нокдауна .
Морфолиновые олигонуклеотиды следует использовать с осторожностью из-за их потенциальных эффектов вне белка-мишени. Для проверки того, является ли наблюдаемый фенотип морфанта результатом нокдауна задуманного гена или же он вызван взаимодействием с РНК, не являющейся непосредственной мишенью введенного олигонуклеотида, можно провести второй эксперимент. Этот эксперимент заключается в повторении фенотипа ( фенокопировании ) морфанта с использованием другого морфолинового олигонуклеотида, нацеленного на ту же мишень, но не перекрывающегося с первым олигонуклеотидом .
Доставка
Для того, чтобы морфолиновый олигонуклеотид мог подействовать, он должен попасть в цитозоль клетки и преодолеть клеточную мембрану . После попадания в цитозоль он начинает свободно диффундировать между цитозолем и ядром , как было показано в экспериментах, при которых введение морфолинового нуклеотида в цитозоль клетки изменяло сплайсинг в ядре . Для доставки морфолинового олигонуклеотида в зародыши, клетки культуры или клетки взрослых животных используются различные методы.
Для доставки в зародыши, как правило, используют , причём введение олигонуклеотидов обычно осуществляют на стадии одной или нескольких клеток . Альтернативный метод для доставки морфолиновых олигонуклеотидов в эмбрионы — электропорация, которая позволяет доставить их в ткани на более поздних стадиях развития зародыша .
Наиболее распространённые методы для доставки морфолиновых олигонуклеотидов в клетки культуры — использование эндопортерного пептида ( англ. Endo-Porter peptide ), который вызывает высвобождение морфолиновых олигонуклеотидов из эндосом ; система специальной доставки ( англ. Special Delivery system ), использующая гетеродуплекс ДНК с морфолиновым олигонуклеотидом и в качестве реагента доставки (больше коммерчески недоступна) ; электропорация ; метод соскоба-загрузки ( англ. scrape loading ) . В 2015 году была описана транс-активирующая амфипатическая система доставки, основанная на ДНК, которая предназначена для удобной доставки незаряженных нуклеиновых кислот, имеющих поли(А)-хвост, таких как пептидо-нуклеиновые кислоты и морфолиновые олигонуклеотиды .
Доставка морфолиновых олигонуклеотидов в ткани взрослых животных затруднительна, хотя разработано несколько систем, позволяющих доставить в ткани немодифицированные морфолиновые олигонуклеотиды (в частности, в от природы слабые мышечные клетки при мышечной дистрофии Дюшена или в клетки сосудистого эндотелия , подвергающиеся стрессу при ). Хотя морфолиновые олигонуклеотиды эффективно проходят через межклеточные промежутки в тканях, доставка их в клетки сложна. Системная доставка в клетки многих типов может быть осуществлена при помощи морфолиновых олигонуклеотидов, ковалентно связанных с ( англ. Cell-penetrating peptide ), и, в то время как токсичный эффект оказывают средние дозы связанных пептидов , в условиях in vivo удалось добиться эффективной доставки морфолиновых олигонуклеотидов при дозах пептидов ниже токсичных . Октагуанидиновый дендример , прикреплённый к концу морфолинового олигонуклеотида (т. н. Vivo-Morpholino), обеспечивает его доставку из крови в цитозоль при систематическом введении во взрослого животного .
Медицинское значение
Морфолиновые олигонуклеотиды, способные к эффективной доставке (такие как связанные с пептидами морфолиновые олигонуклеотиды и Vivo-Morpholino), могут быть перспективными средствами для лечения вирусных и генетических заболеваний . Например, Inc. уже разрабатывает морфолиновые олигонуклеотиды — потенциальные лекарства под названием NeuGene. В настоящее время проходят клинические испытания морфолиновые олигонуклеотиды, которые могут использоваться для лечении миодистрофии Дюшенна у человека .
Примечания
- , с. 206.
- П. В. Золотухин, Ю. А. Лебедева, О. Н. Кузьминова, Е. К. Брюханова. // Валеология. — 2013. — № 2 . — С. 27—33 . — ISSN . 5 марта 2016 года.
- Н. М. Белоногова. // Вавиловский журнал генетики и селекции. — 2014. — Т. 18 , № 1 . — С. 147—157 .
- Т. В. Абрамова, В. Н. Сильников. // Усп. хим.. — 2011. — Т. 80 , № 5 . — С. 452—476 .
- ↑ Summerton J. (англ.) // Biochimica et biophysica acta. — 1999. — Vol. 1489, no. 1 . — P. 141—158. — .
- ↑ Draper B. W. , Morcos P. A. , Kimmel C. B. (англ.) // Genesis (New York, N.Y. : 2000). — 2001. — Vol. 30, no. 3 . — P. 154—156. — .
- Heasman J. (англ.) // Developmental biology. — 2002. — Vol. 243, no. 2 . — P. 209—214. — doi : . — .
- Geller B. L. (англ.) // Current opinion in molecular therapeutics. — 2005. — Vol. 7, no. 2 . — P. 109—113. — .
- ↑ Deas T. S. , Bennett C. J. , Jones S. A. , Tilgner M. , Ren P. , Behr M. J. , Stein D. A. , Iversen P. L. , Kramer L. D. , Bernard K. A. , Shi P. Y. (англ.) // Antimicrobial agents and chemotherapy. — 2007. — Vol. 51, no. 7 . — P. 2470—2482. — doi : . — .
- McClorey G. , Fall A. M. , Moulton H. M. , Iversen P. L. , Rasko J. E. , Ryan M. , Fletcher S. , Wilton S. D. (англ.) // Neuromuscular disorders : NMD. — 2006. — Vol. 16, no. 9-10 . — P. 583—590. — doi : . — .
- ↑ Summerton J. , Weller D. (англ.) // Antisense & nucleic acid drug development. — 1997. — Vol. 7, no. 3 . — P. 187—195. — .
- Nasevicius A. , Ekker S. C. (англ.) // Nature genetics. — 2000. — Vol. 26, no. 2 . — P. 216—220. — doi : . — .
- Heasman J. , Kofron M. , Wylie C. (англ.) // Developmental biology. — 2000. — Vol. 222, no. 1 . — P. 124—134. — doi : . — .
- Howard E. W. , Newman L. A. , Oleksyn D. W. , Angerer R. C. , Angerer L. M. (англ.) // Development (Cambridge, England). — 2001. — Vol. 128, no. 3 . — P. 365—375. — .
- Kos R. , Reedy M. V. , Johnson R. L. , Erickson C. A. (англ.) // Development (Cambridge, England). — 2001. — Vol. 128, no. 8 . — P. 1467—1479. — .
- ↑ Morcos P. A. (англ.) // Genesis (New York, N.Y. : 2000). — 2001. — Vol. 30, no. 3 . — P. 94—102. — .
- ↑ Summerton J. E. (англ.) // Annals of the New York Academy of Sciences. — 2005. — Vol. 1058. — P. 62—75. — doi : . — .
- Stancheva I. , Collins A. L. , Van den Veyver I. B. , Zoghbi H. , Meehan R. R. (англ.) // Molecular cell. — 2003. — Vol. 12, no. 2 . — P. 425—435. — .
- Bruno I. G. , Jin W. , Cote G. J. (англ.) // Human molecular genetics. — 2004. — Vol. 13, no. 20 . — P. 2409—2420. — doi : . — .
- Vetrini F. , Tammaro R. , Bondanza S. , Surace E. M. , Auricchio A. , De Luca M. , Ballabio A. , Marigo V. (англ.) // Human mutation. — 2006. — Vol. 27, no. 5 . — P. 420—426. — doi : . — .
- Morcos P. A. (англ.) // Biochemical and biophysical research communications. — 2007. — Vol. 358, no. 2 . — P. 521—527. — doi : . — .
- König H. , Matter N. , Bader R. , Thiele W. , Müller F. (англ.) // Cell. — 2007. — Vol. 131, no. 4 . — P. 718—729. — doi : . — .
- Kloosterman W. P. , Wienholds E. , Ketting R. F. , Plasterk R. H. (англ.) // Nucleic acids research. — 2004. — Vol. 32, no. 21 . — P. 6284—6291. — doi : . — .
- Flynt A. S. , Li N. , Thatcher E. J. , Solnica-Krezel L. , Patton J. G. (англ.) // Nature genetics. — 2007. — Vol. 39, no. 2 . — P. 259—263. — doi : . — .
- Kloosterman W. P. , Lagendijk A. K. , Ketting R. F. , Moulton J. D. , Plasterk R. H. (англ.) // Public Library of Science Biology. — 2007. — Vol. 5, no. 8 . — P. e203. — doi : . — .
- Lagendijk A. K. , Moulton J. D. , Bakkers J. (англ.) // Biology open. — 2012. — Vol. 1, no. 6 . — P. 566—569. — doi : . — .
- Yen L. , Svendsen J. , Lee J. S. , Gray J. T. , Magnier M. , Baba T. , D'Amato R. J. , Mulligan R. C. (англ.) // Nature. — 2004. — Vol. 431, no. 7007 . — P. 471—476. — doi : . — .
- Matter N. , König H. (англ.) // Nucleic acids research. — 2005. — Vol. 33, no. 4 . — P. e41. — doi : . — .
- Howard M. T. , Gesteland R. F. , Atkins J. F. (англ.) // RNA (New York, N.Y.). — 2004. — Vol. 10, no. 10 . — P. 1653—1661. — doi : . — .
- Penn A. C. , Balik A. , Greger I. H. (англ.) // Nucleic acids research. — 2013. — Vol. 41, no. 2 . — P. 1113—1123. — doi : . — .
- ↑ Bill B. R. , Petzold A. M. , Clark K. J. , Schimmenti L. A. , Ekker S. C. (англ.) // Zebrafish. — 2009. — Vol. 6, no. 1 . — P. 69—77. — doi : . — .
- Kamachi Y. , Okuda Y. , Kondoh H. (англ.) // Genesis (New York, N.Y. : 2000). — 2008. — Vol. 46, no. 1 . — P. 1—7. — doi : . — .
- Hudziak R. M. , Barofsky E. , Barofsky D. F. , Weller D. L. , Huang S. B. , Weller D. D. (англ.) // Antisense & nucleic acid drug development. — 1996. — Vol. 6, no. 4 . — P. 267—272. — .
- Youngblood D. S. , Hatlevig S. A. , Hassinger J. N. , Iversen P. L. , Moulton H. M. (англ.) // Bioconjugate chemistry. — 2007. — Vol. 18, no. 1 . — P. 50—60. — doi : . — .
- Jon D. Moulton. A Brief Introduction to Morpholino Antisense. — 2011.
- Ekker S. C. , Larson J. D. (англ.) // Genesis (New York, N.Y. : 2000). — 2001. — Vol. 30, no. 3 . — P. 89—93. — .
- Robu M. E. , Larson J. D. , Nasevicius A. , Beiraghi S. , Brenner C. , Farber S. A. , Ekker S. C. (англ.) // PLoS genetics. — 2007. — Vol. 3, no. 5 . — P. e78. — doi : . — .
- Rosen J. N. , Sweeney M. F. , Mably J. D. (англ.) // Journal of visualized experiments : JoVE. — 2009. — No. 25 . — doi : . — .
- Cerda G. A. , Thomas J. E. , Allende M. L. , Karlstrom R. O. , Palma V. (англ.) // Methods (San Diego, Calif.). — 2006. — Vol. 39, no. 3 . — P. 207—211. — doi : . — .
- Jubin R. (англ.) // Methods in molecular medicine. — 2005. — Vol. 106. — P. 309—322. — .
- Partridge M. , Vincent A. , Matthews P. , Puma J. , Stein D. , Summerton J. (англ.) // Antisense & nucleic acid drug development. — 1996. — Vol. 6, no. 3 . — P. 169—175. — .
- H. V. Jain, D. Verthelyi and S. L. Beaucage. Amphipathic trans-acting phosphorothioate DNA elements mediate the delivery of uncharged nucleic acid sequences in mammalian cells // RSC Adv.. — 2015. — Vol. 5. — P. 65245—65254. — doi : .
- Fletcher S. , Honeyman K. , Fall A. M. , Harding P. L. , Johnsen R. D. , Wilton S. D. (англ.) // The journal of gene medicine. — 2006. — Vol. 8, no. 2 . — P. 207—216. — doi : . — .
- Kipshidze N. N. , Kim H. S. , Iversen P. , Yazdi H. A. , Bhargava B. , New G. , Mehran R. , Tio F. , Haudenschild C. , Dangas G. , Stone G. W. , Iyer S. , Roubin G. S. , Leon M. B. , Moses J. W. (англ.) // Journal of the American College of Cardiology. — 2002. — Vol. 39, no. 10 . — P. 1686—1691. — .
- Abes S. , Moulton H. M. , Clair P. , Prevot P. , Youngblood D. S. , Wu R. P. , Iversen P. L. , Lebleu B. (англ.) // Journal of controlled release : official journal of the Controlled Release Society. — 2006. — Vol. 116, no. 3 . — P. 304—313. — doi : . — .
- Burrer R. , Neuman B. W. , Ting J. P. , Stein D. A. , Moulton H. M. , Iversen P. L. , Kuhn P. , Buchmeier M. J. (англ.) // Journal of virology. — 2007. — Vol. 81, no. 11 . — P. 5637—5648. — doi : . — .
- Amantana A. , Moulton H. M. , Cate M. L. , Reddy M. T. , Whitehead T. , Hassinger J. N. , Youngblood D. S. , Iversen P. L. (англ.) // Bioconjugate chemistry. — 2007. — Vol. 18, no. 4 . — P. 1325—1331. — doi : . — .
- Li Y. F. , Morcos P. A. (англ.) // Bioconjugate chemistry. — 2008. — Vol. 19, no. 7 . — P. 1464—1470. — doi : . — .
- Morcos P. A. , Li Y. , Jiang S. (англ.) // BioTechniques. — 2008. — Vol. 45, no. 6 . — P. 613—614. — .
- Moulton J. D. , Jiang S. (англ.) // Molecules (Basel, Switzerland). — 2009. — Vol. 14, no. 3 . — P. 1304—1323. — doi : . — .
- . Дата обращения: 25 октября 2015. 8 декабря 2015 года.
Литература
- Б. Аппель и др. / под ред. С. Мюллер. — М. : БИНОМ. Лаборатория знаний, 2013. — С. . — 413 с. — ISBN 978-5-9963-0376-2 .
- Subbotina E. , Koganti S. R. , Hodgson-Zingman D. , Zingman L. V. (англ.) // Clinical pharmacology and therapeutics. — 2015. — doi : . — .
- Bhadra J. , Pattanayak S. , Sinha S. (англ.) // Current protocols in nucleic acid chemistry / edited by Serge L. Beaucage ... [et al.]. — 2015. — Vol. 62. — P. 4—65. — doi : . — .
- Moulton H. M. (англ.) // Current pharmaceutical design. — 2013. — Vol. 19, no. 16 . — P. 2963—2969. — .
Ссылки
- .
|
Типы
нуклеиновых кислот
|
||||
|---|---|---|---|---|
| Азотистые основания | ||||
| Нуклеозиды | ||||
| Нуклеотиды | ||||
|
РНК
|
||||
|
ДНК
|
||||
| Аналоги | ||||
| Типы векторов | ||||
|
||||
- 2021-10-18
- 1