RHD (ген)
- 1 year ago
- 0
- 0


Ген ( др.-греч. γένος — род ) — в классической генетике — наследственный фактор, который несёт информацию об определённом признаке или функции организма, и который является структурной и функциональной единицей наследственности . В таком качестве термин «ген» был введён в 1909 году датским ботаником , физиологом растений и генетиком Вильгельмом Йоханнсеном .
После открытия нуклеиновых кислот в качестве носителя наследственной информации определение гена изменилось, и ген стали определять как участок ДНК (у некоторых вирусов — участок РНК ), задающий последовательность мономеров в полипептиде либо в функциональной РНК .
По мере накопления сведений о строении и работе генов определение понятия «ген» продолжало изменяться, однако в настоящее время не существует универсального определения гена, которое удовлетворило бы всех исследователей . Одно из современных определений гена звучит следующим образом: ген представляет собой последовательность ДНК, составляющие сегменты которой не обязательно должны быть физически смежными. Эта последовательность ДНК содержит информацию об одном или нескольких продуктах в виде белка или РНК. Продукты гена функционируют в составе генетических регуляторных сетей, результат работы которых реализуется на уровне фенотипа .
Совокупность генов организма составляют генотип . Генотип наряду с факторами окружающей среды и развитием определяют, каким будет фенотип . Передача генов потомству является основой наследования фенотипических признаков. Большинство биологических признаков является полигенным, то есть находится под влиянием многих генов. Гены могут изменяться в результате мутаций , изменяющих последовательность ДНК. Вследствие мутаций в популяции гены существуют в различных вариантах, называемых аллелями . Разные аллели гена могут кодировать различающиеся версии белка, что может проявляться фенотипически . Гены наряду с участками ДНК, не содержащими генов, входят в состав генома , представляющего собой весь наследственный материал организма.

Экспериментальные доказательства наличия дискретных факторов наследственности впервые были представлены в 1865 году Грегором Менделем в докладе на заседании Общества естествоиспытателей в Брно . В 1866 году доклад был опубликован в печатном виде . Грегор Мендель изучал наследование признаков у гороха , количественно отслеживая частоту признаков у родительских растений и у потомства. В скрещиваниях растений с различными признаками он проанализировал более 8000 растений. В этих экспериментах Мендель продемонстрировал независимое наследование признаков , различие между доминантными и рецессивными признаками, различие между гетерозиготами и гомозиготами , а также явление прерывистого наследования . Результаты своих экспериментов он описал математически и интерпретировал их, предположив, что существуют дискретные, несмешиваемые в потомстве, факторы наследственности.
До работы Менделя доминирующей концепцией в объяснении закономерностей наследования была концепция, которая предполагала, что признаки родителей у потомков смешиваются аналогично смешиванию жидкостей. Этой концепции следует теория пангенезиса, разработанная Чарльзом Дарвином в 1868 году, два года спустя после публикации результатов Менделя . В этой теории Дарвин предположил существование очень мелких частиц, названных им «геммулами», которые смешиваются во время зачатия.
Статья Менделя осталась практически незамеченной после её публикации в 1866 году, но она получила второе «рождение» в конце XIX века, благодаря Хуго де Фризу , Карлу Корренсу и Эриху фон Чермаку , которые пришли к аналогичным выводам в своих собственных исследованиях . В частности, в 1889 году Хуго де Фриз опубликовал свою книгу «Intracellular Pangenesis» , в которой он постулировал, что разные признаки имеют собственные наследственные носители, и что наследование специфических черт у организмов происходит при помощи частиц. Де Фрис назвал эти единицы «пангенами» (Pangens на немецком языке), использовав часть названия теории пангенеза Дарвина.
В 1909 году Вильгельм Йоханнсен ввел термин «ген» , а Уильям Бейтсон — термин « генетика » , в то время как Эдуард Страсбургер все ещё использовал термин «панген» для обозначения основной физической и функциональной единицы наследственности .
Эксперименты, проведённые в 40-е годы американскими бактериологами из Рокфеллеровского института под руководством О. Эвери , показали, что молекулярным хранилищем генетической информации является ДНК. В работах по генетической трансформации пневмококков было показано, что передача признаков от одних бактерий к другим происходит при помощи только одного вещества — ДНК. Ни белок, ни другие химические компоненты клетки этим свойством не обладали . В 1953 году при помощи рентгеновской кристаллографии Розалинд Франклин и Морис Уилкинс получили высококачественные снимки структуры ДНК. Эти снимки помогли Джеймсу Д. Уотсону и Фрэнсису Крику создать модель молекулы двуцепочечной спирали ДНК и сформулировать гипотезу механизма генетической репликации .
В начале 1950-х годов преобладало мнение, что гены в хромосоме действуют как отдельные объекты, неразделимые путем рекомбинации и расположенные как бусы на веревочке. Эксперименты Сеймура Бензера с использованием мутантов , дефектных бактериофагов в области (1955—1959), показали, что отдельные гены имеют простую линейную структуру и, вероятно, эквивалентны линейному сечению ДНК .
В совокупности этот объём исследований установил центральную догму молекулярной биологии , которая утверждает, что белки транслируются с РНК , которая транскрибируется с ДНК . Эта догма с тех пор, как было показано, имеет исключения, такие как обратная транскрипция в ретровирусах . Современное исследование генетики на уровне ДНК известно как молекулярная генетика .
В 1972 году Уолтер Файерс и его команда первыми определили последовательность гена: последовательность белка оболочки . Последующее развитие секвенирования ДНК с методом Сэнгера в 1977 году Фредериком Сангером улучшило эффективность секвенирования и превратило его в рутинный лабораторный инструмент . Автоматизированная версия метода Сангера использовалась на ранних этапах проекта «Геном человека» .
Теории, разработанные в начале XX века для интеграции менделевской генетики с дарвиновской эволюцией , называются современным синтезом , термином, введенным Джулианом Хаксли .
Эволюционные биологи впоследствии модифицировали эту концепцию, такую как Джорджа Уильямса на эволюцию. Он предложил эволюционную концепцию гена как естественного отбора с определением: «то, что разделяет и рекомбинирует с заметной частотой» :24 . С этой точки зрения, молекулярный ген транскрибируется как единое целое, а эволюционный ген наследуется как единое целое. Связанные идеи, подчеркивающие центральную роль генов в эволюции, были популяризированы Ричардом Докинзом .

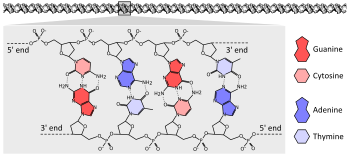
Генетическая информация у подавляющего большинства организмов закодирована в длинных молекулах ДНК . ДНК состоит из двух спирально закрученных полимерных цепей, мономерами которых служат четыре нуклеотида : dАМФ, dГМФ, dЦМФ и dТМФ. Нуклеотиды в ДНК состоят из пятиуглеродного сахара ( 2-дезоксирибозы ), фосфатной группы и одного из четырёх азотистых оснований : аденина , цитозина , гуанина и тимина :2.1 . Азотистое основание связано гликозидной связью с пятиуглеродным (пентозного) сахаром в 1'-положении. Остовом цепей ДНК служит чередующаяся последовательность пентозных сахаров и фосфатов, фосфатные группы присоединяются к сахару в 5'- и 3'-положениях. Номера позиций пентозного кольца отмечены штрихом для того, чтобы различать нумерацию колец в сахаре и азотистом основании .
Из-за химического состава пентозных остатков цепи ДНК имеют направленность. Один конец полимера ДНК содержит открытую гидроксильную группу на дезоксирибозе в 3'-положении; этот конец называется 3'-конец. Другой конец содержит открытую фосфатную группу, это 5'-конец. Две цепи (нити) двойной спирали ДНК ориентированы в противоположных направлениях. Синтез ДНК, в том числе при репликации ДНК , происходит в направлении 5 '→ 3', потому что новые нуклеотиды добавляются посредством реакции дегидратации , которая использует открытый 3’-гидроксил в качестве нуклеофила :27.2 .
Экспрессия генов , закодированных в ДНК, начинается с транскрипции нуклеотидной последовательности ДНК в последовательность нуклеотидов другого типа нуклеиновых кислот — РНК . РНК очень похожа на ДНК, но её мономеры содержат рибозу , а не дезоксирибозу . Кроме того, вместо тимина в РНК используется урацил . Молекулы РНК являются одноцепочечными и менее стабильны, чем ДНК. Гены в ДНК, а после транскрипции и в тРНК, представляют собой природный шифр (код) для синтеза белка. Каждые три нуклеотида (триплет) кодируют одну аминокислоту. К примеру если в гене 300 нуклеотидов, то в белке должно быть 100 аминокислот. Поэтому код называется триплетный. Правило, по которому определяется, какому триплету соответствует какая аминокислота, называется генетическим кодом . Считывание генетического кода происходит в рибосоме во время трансляции РНК в белок . Генетический код почти одинаков для всех известных организмов :4.1 .

Наследственный материал организма, или геном , хранится в одной или нескольких хромосомах , число которых специфично для вида . Хромосома состоит из одной очень длинной молекулы ДНК, которая может содержать тысячи генов :4.2 . Область хромосомы, где находится ген, называется локусом . Каждый локус содержит определённый аллель гена. Представители популяции могут отличаться по аллелям гена, находящимся в одинаковых локусах хромосом.
Большинство эукариотических генов хранятся в нескольких линейных хромосомах. Хромосомы упакованы в ядре в комплексе с белками хроматина . Наиболее многочисленными белками хроматина являются гистоны , которые формируют белковую глобулу, называемую нуклеосомой . ДНК обвивается вокруг нуклеосом, что представляет собой первый уровень упаковки ДНК в хромосоме :4.2 . Распределение нуклеосом вдоль ДНК, а также химические модификации самих гистонов регулируют доступность ДНК для регуляторных факторов, участвующих в репликации, транскрипции, трансляции и репарации. Помимо генов эукариотические хромосомы содержат также служебные последовательности, обеспечивающие стабильность и воспроизведение хромосом, а также их распределение между дочерними клетками в митозе. Это теломеры , сайты инициации репликации и центромера , соответственно :4.2 .
Трудно точно определить, в какую часть последовательности ДНК входит ген .
В молекулярной биологии установлено, что гены — это участки ДНК , несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК , которые определяют развитие, рост и функционирование организма .
Каждый ген характеризуется рядом специфических , таких как промоторы , которые принимают участие в регулировании функции гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания , кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемые cis -регуляторные элементы , англ. cis-regulatory elements ), так и на расстоянии многих миллионов пар оснований ( нуклеотидов ), как в случае с энхансерами , инсуляторами и супрессорами (иногда классифицируемые как trans -регуляторные элементы , англ. trans-regulatory elements ). То есть, понятие гена не ограничено только кодирующим участком ДНК. Ген представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.
Изначально термин «ген» появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Считалось, что такими носителями могут быть только белки , так как их строение (20 аминокислот ) позволяет создать больше вариантов, чем ДНК , которая состоит всего из четырёх видов нуклеотидов . Однако экспериментально доказано, что именно ДНК содержит в себе наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии : ДНК — РНК — белок.
Гены могут подвергаться мутациям — случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК . Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК . Результатом может стать изменённое или даже анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание , или летальными на эмбриональном уровне. Далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода ) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий ( англ. copy number variations ), такими как делеции и дупликации , которые составляют около 1 % всей нуклеотидной последовательности человека . Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена.
Репликация ДНК по большей части чрезвычайно точна, однако ошибки ( мутации ) случаются :7.6 . Частота ошибок в эукариотических клетках может составлять всего 10 −8 в нуклеотиде на репликацию , тогда как для некоторых РНК-вирусов она может достигать 10 −3 . Это означает, что в каждое поколение, каждый человек в геноме накапливает около 30 новых мутаций . Небольшие мутации могут быть вызваны репликацией ДНК и последствиями повреждения ДНК и включают точечные мутации , в которых изменяется одно основание, и мутации со сдвигом рамки , в которых одно основание вставляется или удаляется. Любая из этих мутаций может изменить ген по миссенс (изменить код для кодирования другой аминокислоты) или по нонсенс (преждевременный стоп-кодон ) . Большие мутации могут быть вызваны ошибками в рекомбинации, чтобы вызвать хромосомные аномалии , включая дублирование , делецию, перегруппировку или инверсию больших участков хромосомы. Кроме того, механизмы восстановления ДНК могут вносить мутационные ошибки при восстановлении физического повреждения молекулы. Восстановление, даже с мутацией, является более важным для выживания, чем восстановление точной копии, например, при восстановлении двухцепочечных разрывов :5.4 .
Когда в популяции вида присутствует несколько различных аллелей гена, это называется полиморфизм . Большинство различных аллелей функционально эквивалентны, однако некоторые аллели могут вызывать различные . Самый распространенный аллель гена называется диким типом , а редкие аллели — мутантами . Генетические различия в относительных частотах различных аллелей в популяции обусловлены как естественным отбором , так и генетическим дрейфом . Аллель дикого типа не обязательно является предком менее распространенных аллелей и не обязательно более приспособлена .
и количество генов, которые он содержит, значительно варьируют у таксономических групп. Наименьший геном встречаются у вирусов , и вироидов (которые действуют как один некодирующий ген РНК) . И наоборот, растения могут иметь очень большие геномы , в рисе содержатся более 46 000 генов, кодирующих белок . Общее количество кодирующих белок генов ( протеома Земли), которое оценивалось в 2007 году в 5 млн. последовательностей , к 2017 году было снижено до 3,75 млн .
Генная инженерия — это методы модификации генетического материала для изменения свойств живого организма. С 1970-х годов было разработано , специально предназначенных для добавления, удаления и редактирования генов в вирусах, бактериях, растениях, грибах и животных, включая человека . Недавно разработанные методы геномной инженерии используют инженерные нуклеазные ферменты для создания целевой репарации ДНК в хромосоме , чтобы либо разрушить, либо отредактировать ген в процессе репарации искусственно внесённого разрыва ДНК . Связанный термин синтетическая биология иногда используется для обозначения обширной дисциплины генной инженерии организма .
Генная инженерия в настоящее время является рутинным инструментом при работе с модельными организмами . Например, гены легко добавляются к бактериям , а линии « » мышей с нарушенной функцией определённого гена используются для исследования функции этого гена . Многие организмы были генетически модифицированы для применения в сельском хозяйстве, промышленной биотехнологии, клеточной биологии и медицине .
У многоклеточных организмов обычно модифицируется эмбрион , который вырастает во взрослый генетически модифицированный организм . Однако геномы клеток взрослого организма можно редактировать с использованием методов генной терапии для лечения генетических заболеваний.
| Ключевые понятия |

|
|
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||