Interested Article - Вторичные посредники
- 2021-07-23
- 1
Вторичные посредники , или « вторичные мессенджеры » — это внутриклеточные сигнальные молекулы, высвобождаемые в ответ на стимуляцию рецепторов и вызывающие активацию первичных эффекторных белков . Они же, в свою очередь, приводят к активации вторичных эффекторных белков. Это запускает каскад физиологических изменений, которые могут быть важны для обеспечения таких важных физиологических процессов, как рост, развитие и дифференцировка клеток, активация деления клетки , транскрипция или, наоборот, угнетение транскрипции генов , биосинтез белков, секреция гормонов , нейромедиаторов или цитокинов соответственно типу клетки, изменение биоэлектрической активности клетки, миграция клеток, обеспечение их выживаемости или, наоборот, индукция апоптоза . Вторичные посредники являются инициирующими элементами во множестве внутриклеточных сигнальных каскадов. Вследствие всего этого вторичные посредники играют очень важную роль в жизни клетки, а грубое нарушение работы любой из систем вторичных посредников оказывает неблагоприятное воздействие на клетку (например, может привести к её опухолевой трансформации или, наоборот, к апоптозу).
Вторичные посредники, как правило, являются малыми небелковыми молекулами. Важнейшие примеры молекул вторичных посредников (но не ограничивающиеся ими) включают в себя циклический АМФ , циклический ГМФ , инозитолтрифосфат , диацилглицерин , кальций , оксид азота (II) . Клетка выделяет (или, наоборот, уменьшает выделение) тех или иных вторичных посредников в ответ на воздействие внеклеточных сигнальных молекул — так называемых «первичных сигнальных молекул» или «первичных мессенджеров». Первичными сигналами могут являться, например, гормоны (пептидные, моноаминовые и другие), нейромедиаторы, цитокины и др. Поскольку первичные мессенджеры обычно являются высоко гидрофильными молекулами (как, например, адреналин или серотонин ), или достаточно крупными полипептидными молекулами (как окситоцин и инсулин ), они не могут преодолеть фосфолипидный бислой мембраны клетки, для того, чтобы непосредственно инициировать каскад внутриклеточных физиологических изменений. Это функциональное ограничение и вынудило клетки эволюционным путём приобрести механизмы трансмембранной передачи внеклеточных сигналов внутрь клетки, основывающиеся на существовании цепочки «рецептор для внеклеточного сигнала -> первичный эффектор -> вторичный посредник (вторичный мессенджер) -> вторичный эффектор -> дальнейшие сигнальные каскады». Важной возможностью, встроенной в эту систему передачи сигнала, является то, что на каждом этапе передачи сигнала внутрь клетки первичный сигнал подвергается многократному усилению, амплификации (одна молекула первичного сигнала и, соответственно, активация одного первичного эффектора приводит к высвобождению множества молекул вторичного посредника, такого, как цАМФ, а множество молекул мультиспецифичных и мультицикличных вторичных эффекторов, таких, как протеинкиназа A , активированных при участии этого вторичного посредника, приводят к фосфорилированию и, соответственно, угнетению или активации ещё большего множества разнообразных белков). Другой пример — сигнал Ras . GTP связывается с MAP-киназой , что приводит к аллостерической активации целого ряда факторов пролиферации и индукции митоза, таких, как Myc и CREB .
Эрл Сазерленд впервые открыл систему вторичных посредников, за что получил в 1971 году Нобелевскую премию по физиологии и медицине. Эрл Сазерленд заметил, что адреналин стимулирует печень к образованию глюкозы из гликогена (процесс, называемый гликогенолизом ), однако это происходит не непосредственно при воздействии адреналина на клетку, а при посредничестве цАМФ (то есть, одного лишь действия адреналина на клетку недостаточно для того, чтобы запустить процесс глюконеогенеза — для этого сначала должен произойти ряд внутриклеточных событий, а для того, чтобы они могли произойти, в клетке должен выполняться ряд предварительных условий для инициирования каскада передачи сигнала, в частности, иметься доступная, не заблокированная, аденилатциклаза). Сазерленд показал, что адреналин вызывает сначала увеличение концентрации внутриклеточного цАМФ, а уже затем (в результате запуска каскада внутриклеточных событий) образование глюкозы из гликогена. Он также смог показать, что если заблокировать аденилатциклазу и тем самым предотвратить вызываемое адреналином увеличение концентрации внутриклеточного цАМФ, то адреналин не оказывает действия (не вызывает образования глюкозы из гликогена). Механизм был позже исследован в деталях Мартином Родбеллом и Альфредом Гилманом, которые за это также получили Нобелевскую премию по медицине и физиологии в 1994 году .
Вторичные посредники могут быть синтезированы и активированы при помощи различных ферментов , например, циклаз ( аденилатциклаза , ), которые синтезируют циклические нуклеотиды цАМФ и цГМФ , или синтазы оксида азота , которая синтезирует оксид азота (II) , или фосфолипазы C , которая синтезирует диацилглицерин и инозитолтрифосфат , или же при помощи открытия или закрытия ионных каналов клетки, которые позволяют вход в клетку или выход из клетки тех ионов, как это происходит при передаче сигнала при помощи ионов кальция . Эти малые молекулы вторичных посредников связываются со вторичными эффекторными белками, в частности, активируют протеинкиназы, открывают и закрывают ионные каналы и т. д., что в свою очередь запускает дальнейший внутриклеточный сигнальный каскад вплоть до эффекторов N-го порядка.
Типы молекул вторичных посредников

Существуют три основных типа молекул вторичных посредников:
- Гидрофобные молекулы: нерастворимые в воде молекулы, такие, как диацилглицерин и фосфатидилинозитол, которые связываются с мембранами клетки и диффундируют с цитоплазматической мембраны клетки по межмембранным пространствам на внутренние клеточные мембраны, включая мембрану ядра и ядрышка, где они могут «добраться» и воздействовать на мембранно-связанные вторичные эффекторные белки;
- Гидрофильные молекулы: водорастворимые молекулы, такие, как цАМФ , цГМФ , инозитолтрифосфат , кальций , которые распределяются в водной среде цитоплазмы клетки;
- Газы : оксид азота (II) , оксид углерода (II) , сероводород , которые могут как диффундировать в цитоплазме клетки, так и преодолевать клеточные мембраны.
Общие свойства молекул вторичных посредников
Внутриклеточные вторичные посредники имеют некоторые общие для них всех свойства:
- Они могут быть очень быстро синтезированы и выделены и так же быстро удалены или обезврежены при помощи специфических каталитических ферментов или ионных каналов;
- Некоторые из них, такие, как ионы кальция, могут храниться про запас в специализированных органеллах (гранулах или вакуолях) и быстро высвобождаться — и так же быстро связываться и транспортироваться — при необходимости.
- Их продукция и выделение, как и их удаление, уничтожение или обезвреживание, находится под жёстким контролем внутриклеточных систем обратной связи, не допускающих чрезмерного усиления или чрезмерной продолжительности поступающего извне сигнала и предотвращающих, в нормальных условиях, самоповреждение клетки в процессе обработки сигнала.
- Их продукция и выделение, как и их удаление, уничтожение или обезвреживание, может быть ограничено во времени и пространстве, что позволяет клетке локализовать в пространстве (в той или иной части клетки) и ограничить во времени процессы передачи сигнала.
Общие механизмы систем вторичных посредников

В каждой клетке существует несколько систем вторичных посредников, в частности система циклического АМФ , фосфатидилинозитоловая система (диацилглицерин и инозитолтрифосфат), система арахидоновой кислоты , система оксида азота , система кальция , система циклического ГМФ и другие, однако все они очень похожи по общему устройству механизма, несмотря на то, что вовлечённые в реализацию их эффектов и общие эффекты их активации могут очень различаться.
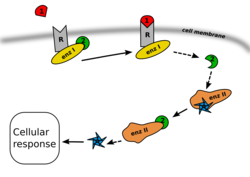
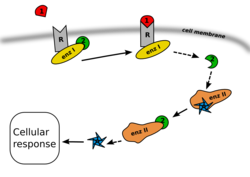
В большинстве случаев всё начинается с того, что тот или иной внеклеточный лиганд-агонист связывается со специфическим сайтом (доменом связывания лиганда) на трансмембранном рецепторном белке. Связывание лиганда с этим сайтом рецептора приводит к изменению его пространственной конфигурации. Это изменение пространственной конфигурации рецептора может привести к изменению его активности и к запуску того или иного первичного эффектора. Результатом действия первичного эффектора является синтез и выделение (или, наоборот, угнетение синтеза) тех или иных вторичных посредников, то есть генерация вторичного сигнала (вторичного месседжа, будь то положительный месседж — синтез вторичного посредника, или отрицательный месседж — угнетение синтеза вторичного посредника).
Например, в случае с G-белок-связанными рецепторами, изменение конформации белка-рецептора, происходящее при связывании с ним лиганда, приводит к тому, что специфический внутриклеточный домен рецептора приобретает способность связываться с G-белком. G-белок называется так по ГДФ и ГТФ , которые этот белок способен связывать. G-белок гидрофобен, поэтому он связан с внутренней мембраной клетки, что облегчает связывание G-белка с трансмембранными рецепторами. G-белок является передатчиком и трансформатором сигнала от рецептора внутрь клетки; он состоит из трёх субъединиц: α, β и γ.
Когда G-белок связывается с активированным рецептором, он приобретает способность к обмену ГДФ (гуанозиндифосфата), связанного с его α-субъединицей, на ГТФ (гуанозинтрифосфат). После того, как этот обмен произойдёт, α-субъединица G-белка приобретает способность диссоциироваться от β- и γ-субъединиц. Все три субъединицы при этом остаются мембранно-связанными. После того как α-субъединица диссоциирует от β- и γ-субъединиц и приобретает способность диффундировать вдоль внутреннего слоя клеточной мембраны, α-субъединица, в конце концов, сталкивается в процессе этого движения с другим мембранным белком — так называемым «первичным эффектором», которым в случае G-белка обычно является аденилатциклаза.
В результате этого взаимодействия α-субъединицы G-белка с первичным эффектором (например, с аденилатциклазой), ферментативная активность первичного эффекторного белка изменяется тем или иным образом — в зависимости от типа α-субъединицы (ингибиторной или стимулирующей). В результате первичный эффекторный белок производит вторичный сигнал, которым является, соответственно, уменьшение или увеличение продукции того или иного вещества из субстрата его ферментативной активности. Для аденилатциклазы субстратом является АТФ , а конечным продуктом реакции — . Для первичного эффектора - фосфолипазы C — субстратом является фосфатидилинозитол , а продуктами реакции — диацилглицерин и инозитолтрифосфат . Для субстратом является L- аргинин , а продуктом реакции — оксид азота (II) . Эти продукты реакций и являются вторичными посредниками (вторичными мессенджерами), в то время как лиганд-агонист рецептора называется первичным сигналом или первичным мессенджером. Далее вторичный посредник (или вторичные посредники, если их несколько) могут диффундировать в цитоплазму клетки или в межмембранные пространства (в зависимости от гидрофильности или липофильности), тем самым эффективно передавая многократно усиленный сигнал внутрь клетки. В клетке эти вторичные посредники, в свою очередь, активируют различные так называемые «вторичные эффекторы» (например, в случае цАМФ это прежде всего , в случае инозитолтрифосфата — прежде всего протеинкиназа C ; мишеней для воздействия может быть несколько или даже много). А вторичные эффекторы запускают дальнейший каскад нисходящих эффекторных реакций. Конечные эффекты зависят как от типа рецептора и G-белка (стимулирующий или ингибирующий), так и от конкретной системы вторичных посредников и вторичных эффекторов.
В то же время β- и γ-субъединицы G-белка способны активировать или угнетать (в зависимости от типа G-белка) добавочные сигнальные пути, например, GIRK (входящие однонаправленные каналы ионов калия) или фосфолипазу C или синтазу оксида азота. Таким образом, один и тот же рецептор способен активировать сразу несколько систем вторичных посредников и производить «кросс-общение» с сигнальными путями, считающимися более характерными для других типов рецепторов.
Ионы кальция являются одним из важных типов вторичных посредников, которые ответственны за осуществление множества важных физиологических функций, таких, как сокращение мышц, оплодотворение, выделение нейромедиаторов. Ионы кальция в норме связаны со специальными белками и/или хранятся в специальных внутриклеточных компартментах (в частности, в гладком эндоплазматическом ретикулуме), откуда могут быть быстро высвобождены в процессе передачи сигнала. Фермент фосфолипаза C (первичный эффектор) продуцирует вторичные посредники диацилглицерин и инозитолтрифосфат, что приводит к повышению проницаемости каналов ионов кальция. Активный G-белок открывает кальциевые каналы клетки, что приводит к входящему току ионов кальция. Другой вторичный посредник, также являющийся продуктом фосфолипазы C, диацилглицерин, активирует протеинкиназу C, которая способствует активации аденилатциклазы и накоплению цАМФ (ещё одного вторичного посредника).
Примеры
Примечания
- Kimball, J. . Архивировано из 7 февраля 2006 года.
- MeSH
- Reece, Jane; Campbell, Neil. Biology (неопр.) . — San Francisco: , 2002. — ISBN 0-8053-6624-5 .
- . Дата обращения: 22 февраля 2015. 23 января 2018 года.
- от 23 января 2018 на Wayback Machine , Nobelprize.org
Ссылки
- Kimball, J. . Дата обращения: 10 февраля 2006. Архивировано из 7 февраля 2006 года.
- 2021-07-23
- 1