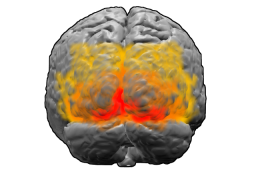
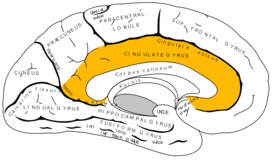
Кора больших полушарий
- 1 year ago
- 0
- 0
Кора́ больши́х полуша́рий головно́го мо́зга или кора́ головно́го мо́зга ( лат. cortex cerebri ) — структура головного мозга , слой серого вещества толщиной 1,3−4,5 мм , расположенный по периферии полушарий большого мозга и покрывающий их. Наибольшая толщина отмечается в верхних участках предцентральной, постцентральной извилин и парацентральной дольки .
Кора головного мозга играет очень важную роль в осуществлении высшей нервной (психической) деятельности .
Кора головного мозга человека составляет более 80 % массы мозга .
У человека кора составляет в среднем 44 % от объёма всего полушария в целом . Площадь поверхности коры одного полушария у взрослого человека 1200 см² (в основном от 1000 до 1400 см²) . На поверхностные части приходится 1 / 3 , на залегающие в глубине между извилинами — 2 / 3 всей площади коры .
Величина и форма борозд подвержены значительным индивидуальным колебаниям — не только мозг различных людей, но даже полушария одной и той же особи по рисунку борозд не вполне похожи .
Всю кору полушарий принято разделять на 4 типа: древняя ( палеокортекс ), старая ( архикортекс ), новая ( неокортекс ) и промежуточная кора (состоящая из промежуточной древней и промежуточной старой коры). Поверхность неокортекса у человека занимает 95,6 %, архикортекса — 2,2 %, палеокортекса — 0,6 %, промежуточной — 1,6 % .
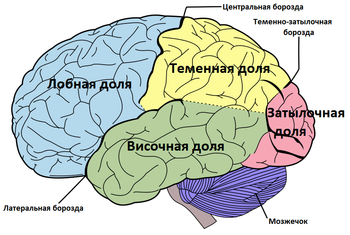
Кора большого мозга покрывает поверхность полушарий и образует большое количество различных по глубине и протяжённости борозд ( лат. sulci cerebri ). Между бороздами расположены различной величины извилины большого мозга ( лат. gyri cerebri ) .
В каждом полушарии различают следующие поверхности:
Эти три поверхности каждого полушария, переходя одна в другую, образуют три края. Верхний край ( лат. margo superior ) разделяет верхнелатеральную и медиальную поверхности. Нижнелатеральный край ( лат. margo inferolateralis ) отделяет верхнелатеральную поверхность от нижней. Нижнемедиальный край ( лат. margo inferomedialis ) располагается между нижней и медиальной поверхностями .
В каждом полушарии различают наиболее выступающие места: спереди — лобный полюс ( лат. polus frontalis ), сзади — затылочный ( лат. polus occipitalis ), и сбоку — височный ( лат. polus temporalis ) .
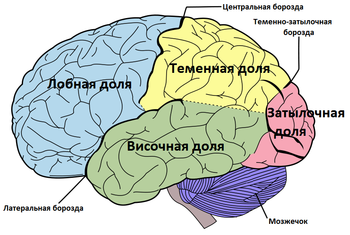
Полушарие разделено на пять долей. Четыре из них примыкают к соответствующим костям свода черепа:
В книге Годфруа Ж. «Что такое психология» выделяется шестая доля, мозолистого тела, расположенная на внутренней стороне полушария под мозолистым телом.
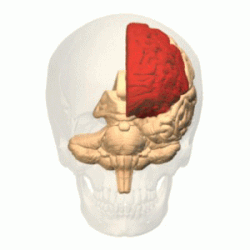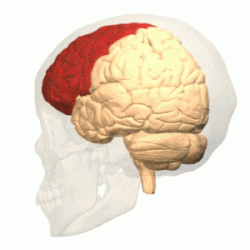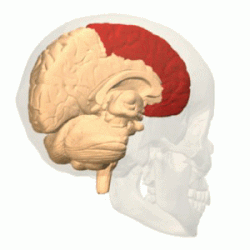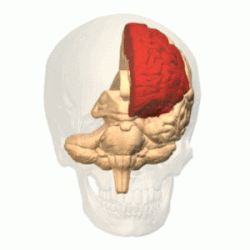
Лобную долю от теменной отделяет глубокая центральная (роландова) борозда ( лат. sulcus centralis ). Она начинается на медиальной поверхности полушария, переходит на его верхнелатеральную поверхность, идёт по ней немного косо, сзади наперёд, и обычно не доходит до латеральной (боковой или сильвиевой) борозды мозга .
Приблизительно параллельно центральной борозде располагается предцентральная борозда ( лат. sulcus precentralis ), которая не доходит до верхнего края полушария. Предцентральная борозда окаймляет спереди прецентральную извилину ( лат. gyrus precentralis ) .
Верхняя и нижняя лобные борозды ( лат. sulci frontales superior et inferior ) направляются от предцентральной борозды вперёд. Они делят лобную долю на:
Латеральная борозда ( лат. sulcus lateralis ) — одна из наиболее глубоких борозд головного мозга. Она отделяет височную долю от лобной и теменной. Залегает латеральная борозда на верхнелатеральной поверхности каждого полушария и идёт сверху вниз и кпереди. В глубине этой борозды располагается углубление — латеральная ямка большого мозга ( лат. fossa lateralis cerebri ), дном которой является наружная поверхность островка .
От латеральной борозды к верху отходят мелкие борозды, называемые ветвями. Наиболее постоянными из них являются восходящая ( лат. ramus ascendens ) и передняя ( лат. ramus anterior ) ветви. Верхнезадний отдел борозды называется задней ветвью ( лат. ramus posterior ) .
Нижняя лобная извилина , в пределах которой проходят восходящая и передняя ветви, разделяется ими на три части:
Залегает сзади от центральной борозды, которая отделяет её от лобной. От височной отграничена латеральной бороздой мозга, от затылочной — частью теменно-затылочной борозды ( лат. sulcus parietooccipitalis ) .
Параллельно прецентральной извилине проходит постцентральная ( лат. gyrus postcentralis ). От неё сзади, почти параллельно продольной щели большого мозга, идёт внутритеменная борозда ( лат. sulcus intraparietalis ), делящая задневерхние отделы теменные отделы теменной доли на две извилины: верхнюю ( лат. lobulus parietalis superior ) и нижнюю ( лат. lobulus parietalis inferior ) теменные дольки. В нижней теменной дольке различают две сравнительно небольшие извилины: надкраевую ( лат. gyrus supramarginalis ), лежащую впереди и замыкающую задние отделы латеральной борозды, и расположенную кзади от предыдущей угловую ( лат. gyrus angularis ), которая замыкает верхнюю височную борозду .
Между восходящей и задней ветвями латеральной борозды мозга расположен участок коры, обозначаемый как лобно-теменная покрышка ( лат. operculum frontoparietalis ). В неё входят задняя часть нижней лобной извилины, нижние отделы предцентральной и постцентральной извилин, а также нижний отдел передней части теменной доли .
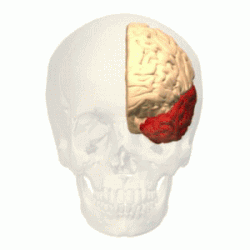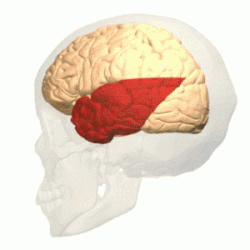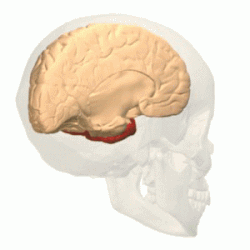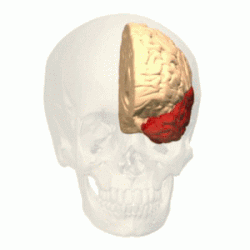
Имеет наиболее выраженные границы. В ней различают выпуклую латеральную поверхность и вогнутую нижнюю. Тупой полюс височной доли обращён вперёд и несколько вниз. Латеральная борозда большого мозга резко отграничивает височную долю от лобной .
Две борозды, расположенные на верхнелатеральной поверхности: верхняя ( лат. sulcus temporalis superior ) и нижняя ( лат. sulcus temporalis inferior ) височные борозды, следуя почти параллельно латеральной борозде мозга, разделяют долю на три височные извилины: верхнюю, среднюю и нижнюю ( лат. gyri temporales superior, medius et inferior ) .
Те участки височной доли, которые направлены в сторону латеральной борозды мозга изрезаны короткими поперечными височными бороздами ( лат. sulci temporales transversi ). Между этими бороздами залегают 2-3 короткие поперечные височные извилины, связанные с извилинами височной доли ( лат. gyri temporales transversi ) и островком .
Залегает на дне латеральной ямки большого мозга ( лат. fossa lateralis cerebri ).
Она представляет собой трёхстороннюю пирамиду, обращённую своей вершиной — полюсом островка — кпереди и кнаружи, в сторону латеральной борозды. С периферии островок окружён лобной, теменной и височной долями, участвующими в образовании стенок латеральной борозды мозга .
Основание островка с трёх сторон окружено круговой бороздой островка ( лат. sulcus circularis insulae ).
Его поверхность прорезана глубокой центральной бороздой островка ( лат. sulcus centralis insulae ). Эта борозда разделяет островок на переднюю и заднюю части .
На поверхности различают большое количество мелких извилин островка ( лат. gyri insulae ). Большая передняя часть состоит из нескольких коротких извилин островка ( лат. gyri breves insulae ), задняя — одной длинной извилины ( лат. gyrus longus insulae ) .
На медиальную поверхность полушария выходят лобная, теменная и затылочная доли.
Поясная извилина ( лат. gyrus cinguli ) начинается подмозолистым полем ( лат. area subcallosa ), огибает мозолистое тело и при посредстве узкой полоски — перешейка поясной извилины ( лат. isthmus gyri cinguli ) переходит в парагиппокампальную извилину на нижней поверхности полушария .
Борозда мозолистого тела ( лат. g sulcus corporis callosi ) отделяет поясную извилину от мозолистого тела и на нижней поверхности полушария продолжается в борозду гиппокампа .
Поясная извилина ограничена сверху поясной бороздой ( лат. sulcus cinguli ). В последней различают выпуклую по направлению к лобному полюсу переднюю часть и заднюю часть, которая, следуя вдоль поясной извилины и не доходя до её заднего отдела, поднимается к верхнему краю полушария большого мозга. Задний конец борозды лежит позади верхнего конца центральной борозды. Между предцентральной бороздой, окончание которой иногда хорошо видно у верхнего края медиальной поверхности полушария, и концом поясной борозды, располагается парацентральная долька ( лат. lobulus paracentralis ) .
Выше поясной извилины, Спереди от подмозолистого поля, начинается медиальная лобная извилина ( лат. gyrus frontalis medialis ). Она тянется до парацентральной дольки и является нижней частью верхней лобной извилины.
Сзади от поясной борозды лежит небольшая четырёхугольная долька — предклинье ( лат. precuneus ). Её задней границей является глубокая теменно-затылочная борозда ( лат. sulcus parietooccipitalis ), нижней — подтеменная борозда ( лат. sulcus subparietalis ), отделяющая предклинье от заднего отдела поясной извилины .
Сзади и ниже предклинья залегает треугольная долька — клин ( лат. cuneus ). Выпуклая наружная поверхность клина участвует в образовании затылочного полюса. Направленная вниз и вперёд вершина клина почти доходит до заднего отдела поясной извилины. Задненижней границей клина является очень глубокая шпорная борозда ( лат. sulcus calcarinus ), передней — теменно-затылочная борозда .
На нижней поверхности лобной доли располагается обонятельная борозда ( лат. sulcus olfactorius ). Кнутри от неё, между нею и нижнемедиальным краем полушария, лежит прямая извилина ( лат. gyrus rectus ). Её задний отдел доходит до переднего продырявленного вещества ( лат. substantia perforata anterior ). Кнаружи от борозды располагается остальная часть нижней поверхности лобной доли, изрезанная короткими глазничными бороздами ( лат. sulci orbitales ), на ряд небольших глазничных извилин ( лат. gyri orbitales ) .
Нижняя поверхность височной доли глубокой бороздой гиппокампа ( лат. sulcus hippocampi ) отделена от ножек мозга. В глубине борозды залегает узкая зубчатая извилина ( лат. gyrus dentatus ). Передний её конец переходит в крючок, а задний — в ленточную извилину ( лат. gyrus fasciolaris ) залегающую под валиком мозолистого тела. Латерально от борозды находится парагиппокампальная извилина ( лат. gyrus parahippocampalis ). Впереди эта извилина имеет утолщение в виде крючка ( лат. uncus ), а кзади продолжается в язычную извилину ( лат. gyrus lingualis ). Парагиппокампальную и язычную извилины с латеральной стороны ограничивает коллатеральная борозда ( лат. sulcus collateralis ), переходящая спереди в носовую борозду ( лат. sulcus rhinalis ). Остальную часть нижней поверхности височной доли занимают медиальная и латеральная затылочно-височные извилины ( лат. gyri occipitotemporales medialis et lateralis ), разделённые затылочно-височной бороздой ( лат. sulcus occipitotemporalis ). Латеральная затылочно-височная извилина нижнелатеральным краем полушария отделяется от нижней височной извилины .
Цитоархитектоника (расположение клеток)
Миелоархитектоника (расположение волокон)
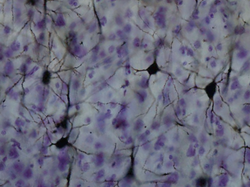
Кора полушарий головного мозга представлена слоем серого вещества толщиной в среднем около 3 мм (1,3 — 4,5 мм). Наиболее сильно развита она в передней центральной извилине. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга. В коре содержится около 10-14 млрд нервных клеток. Различные её участки, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток (цитоархитектоника), расположения волокон (миелоархитектоника) и функциональным значением, называются полями . Они представляют собой места высшего анализа и синтеза нервных импульсов . Резко очерченные границы между ними отсутствуют. Для коры характерно расположение клеток и волокон слоями .
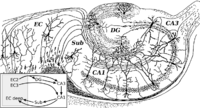
Типичным для новой коры ( лат. neocortex ) является наличие шести слоёв, различающихся между собой главным образом по форме входящих в них нервных клеток. При этом на медиальной и нижней поверхностях полушарий сохранились участки старой ( лат. archipallium ) и древней ( лат. paleopallium ) коры, имеющей 2-слойное и 3-слойное строение . Также выделяется промежуточная кора ( лат. mesopallium ) располагающаяся между старой и новой, а также древней и новой корой . Древняя кора представлена гиппокампом , а старая — участком коры возле обонятельной луковицы на нижней поверхности лобной доли .
Мультиполярные нейроны коры головного мозга весьма разнообразны по форме. Среди них можно выделить:
Пирамидные нейроны составляют основную и наиболее специфическую для коры головного мозга форму (80—90 % всех нейронов). Размеры их варьируют от 10 до 140 мкм. Они имеют вытянутое треугольное тело, вершина которого обращена к поверхности коры. От вершины и боковых поверхностей тела отходят дендриты , заканчивающиеся в различных слоях серого вещества. От основания пирамидных клеток берут начало аксоны , в одних клетках короткие, образующие ветвления в пределах данного участка коры, в других — длинные, поступающие в белое вещество .
Пирамидные клетки различных слоёв коры отличаются размерами и имеют разное функциональное значение. Мелкие клетки представляют собой вставочные нейроны, аксоны которых связывают отдельные участки коры одного полушария (ассоциативные нейроны) или двух полушарий (комиссуральные нейроны). Эти клетки встречаются в разных количествах во всех слоях коры. Особенно богата ими кора головного мозга человека. Аксоны крупных пирамидных нейронов принимают участие в образовании пирамидных путей , проецирующих импульсы в соответствующие центры мозгового ствола и спинного мозга .
Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоёв:
Кора полушарий головного мозга также содержит мощный нейроглиальный аппарат, выполняющий трофическую, защитную, опорную и разграничительную функции .
На медиальной и нижней поверхности полушарий сохранились участки старой, древней коры, которые имеют двухслойное и трехслойное строение.
Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их аксоны проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Основная масса волокон этого сплетения представлена ветвлениями дендритов нейронов нижележащих слоёв .
Наружный зернистый слой образован мелкими нейронами диаметром около 10 мкм, имеющими округлую, угловатую и пирамидальную форму, и звёздчатыми нейронами. Дендриты этих клеток поднимаются в молекулярный слой. Аксоны или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя .
Является самым широким по сравнению с другими слоями коры головного мозга. Он особенно хорошо развит в прецентральной извилине. Величина пирамидных клеток последовательно увеличивается в пределах 10-40 мкм от наружной зоны этого слоя к внутренней. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Дендриты, берущие начало от боковых поверхностей пирамиды и её основания, имеют незначительную длину и образуют синапсы со смежными клетками этого слоя. Аксон пирамидной клетки всегда отходит от её основания. В мелких клетках он остаётся в пределах коры; аксон же, принадлежащий крупной пирамиде, обычно формирует миелиновое ассоцативное или комиссуральное волокно, идущее в белое вещество .
В некоторых полях коры развит очень сильно (например, в зрительной зоне коры ). Однако в других участках он может отсутствовать (в прецентральной извилине). Этот слой образован мелкими звёздчатыми нейронами. В его состав входит большое количество горизонтальных волокон .
Образован крупными пирамидными клетками, причём область прецентральной извилины содержит гигантские клетки, описанные впервые российским анатомом В. А. Бецем в 1874 году ( клетки Беца ). Они достигают в высоту 120 и в ширину 80 мкм. В отличие от других пирамидных клеток коры, гигантские клетки Беца характеризуются наличием крупных глыбок хроматофильного вещества. Их аксоны образуют главную часть кортико-спинальных и кортико-нуклеарных путей и оканчиваются на мотонейронах мозгового ствола и спинного мозга .
Перед выходом из коры от пирамидного пути отходит множество коллатералей. Аксоны от гигантских клеток Беца дают коллатерали, посылающие тормозящие импульсы в саму кору. Также коллатерали волокон пирамидного пути идут в полосатое тело , красное ядро, ретикулярную формацию, ядра моста и нижних олив. Ядра моста и нижних олив передают сигнал в мозжечок . Таким образом, когда пирамидный путь передаёт сигнал, вызывающий целенаправленное движение, в спинной мозг , одновременно сигналы получают базальные ганглии , ствол мозга и мозжечок . Помимо коллатералей пирамидных путей , существуют волокна, которые идут непосредственно от коры к промежуточным ядрам: хвостатому телу, красному ядру, ядрам ретикулярной формации ствола мозга и др.
Образован нейронами различной, преимущественно веретенообразной формы. Внешняя зона этого слоя содержит более крупные клетки. Нейроны внутренней зоны мельче и лежат на большом расстоянии друг от друга. Аксоны клеток полиморфного слоя уходят в белое вещество в составе эфферентных путей головного мозга. Дендриты достигают молекулярного слоя коры .
Среди нервных волокон коры полушарий головного мозга можно выделить:
Кроме тангенциального сплетения молекулярного слоя, на уровне внутреннего зернистого и ганглионарного слоёв расположены два тангенциальных слоя миелиновых нервных волокон и коллатералей аксонов клеток коры. Вступая в синаптические связи с нейронами коры, горизонтальные волокна обеспечивают широкое распространение в ней нервного импульса .
I, II, III, IV, V, VI — слои коры
Афферентные волокна
1. кортико-кортикальное волокно
2. таламо-кортикальное волокно
2а. зона распространения специфических таламо-кортикальных волокон
3.
пирамидные нейроны
3а. заторможенные пирамидные нейроны
4.
тормозные нейроны и их
синапсы
4а. клетки с аксональной кисточкой
4б. малые корзинчатые клетки
4в. большие корзинчатые клетки
4г. аксоаксональные нейроны
4д. клетки с двойным букетом дендритов (тормозящие тормозные нейроны)
5.
шипиковые звёздчатые клетки
, возбуждающие пирамидные нейроны непосредственно и путём стимуляции клеток с двойным букетом дендритов
Исследуя кору больших полушарий головного мозга Я. Сентаготаи и представители его школы установили, что её структурно-функциональной единицей является модуль — вертикальная колонка диаметром около 300 мкм. Модуль организован вокруг кортико-кортикального волокна, представляющего собой аксон пирамидной клетки III слоя (слоя пирамидальных клеток) того же полушария (ассоциативное волокно), либо от пирамидальных клеток противоположного (комиссуральное). В модуль входят два таламо-кортикальных волокна — специфических афферентных волокна, оканчивающихся в IV слое коры на шипиковых звёздчатых нейронах и отходящих от основания (базальных) дендритах пирамидальных нейронов. Каждый модуль, по мнению Сентаготаи разделяется на два микромодуля диаметром менее 100 мкм. Всего в неокортексе человека примерно 3 млн модулей. Аксоны пирамидальных нейронов модуля проецируются на три модуля той же стороны и через мозолистое тело посредством комиссуральных волокон на два модуля противоположного полушария. В отличие от специфических афферентных волокон, оканчивающихся в IV слое коры, кортико-кортикальные волокна образуют окончания во всех слоях коры, и, достигая I слоя, дают горизонтальные ветви, выходящие далеко за пределы модуля .
Помимо специфических (таламо-кортикальных) афферентных волокон, на выходные пирамидальные нейроны возбуждающее влияние оказывают шипиковые звёздчатые нейроны. Различают два типа шипиковых клеток:
Тормозная система модуля представлена следующими типами нейронов:
Система угнетения тормозных нейронов:
Мощный возбуждающий эффект фокальных шипиковых звёздчатых клеток объясняется тем, что они одновременно возбуждают пирамидные нейроны и клетку с двойным букетом дендритов. Таким образом, первые три тормозных нейрона тормозят пирамидные клетки, а клетки с двойным букетом дендритов возбуждают их, угнетая тормозные нейроны .
Однако, также существуют критические и альтернативные концепции , ставящие под сомнение модульную организацию коры больших полушарий и мозжечка. Безусловно, влияние на эти воззрения оказало предсказание в 1985 г. и в дальнейшем открытие в 1992 г. диффузного объёмного нейротрансмиттинга .
Межнейрональные взаимосвязи нейронов коры больших полушарий головного мозга можно представить следующим образом: входящая (афферентная) информация поступает из таламуса по таламо-кортикальным волокнам, которые заканчиваются на клетках IV (внутреннего зернистого) слоя. Его звёздчатые нейроны оказывают возбуждающее воздействие на пирамидные клетки III (пирамидальных нейронов) и V (ганглионарного) слоёв, а также на клетки с двойным букетом дендритов, которые блокируют тормозные нейроны. Клетки III слоя образуют волокна (ассоциативные и комиссуральные), которые связывают между собой различные отделы коры. Клетки V и VI (мультиморфных клеток) слоёв формируют проекционные волокна, которые уходят в белое вещество и несут информацию другим отделам центральной нервной системы. Во всех слоях коры находятся тормозные нейроны, играющие роль фильтра путём блокирования пирамидных нейронов .
Кора различных отделов характеризуется преимущественным развитием тех или иных её слоёв. Так, в двигательных центрах коры, например в передней центральной извилине, сильно развиты III, V и VI и плохо выражены II и IV слои. Это так называемый агранулярный тип коры . Из этих областей берут начало нисходящие проводящие пути центральной нервной системы. В чувствительных корковых центрах, где заканчиваются афферентные проводники, идущие от органов обоняния, слуха и зрения, слабо развиты слои, содержащие крупные и средние пирамидные клетки , тогда как зернистые слои (II и IV) достигают своего максимального развития. Это гранулярный тип коры .
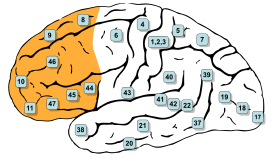
Цитоархитектонические поля Бродмана – отделы коры больших полушарий головного мозга, отличающиеся по своей цитоархитектонике (строению на клеточном уровне). Выделяется 52 цитоархитектонических поля Бродмана.
В 1909 году немецкий невролог Корбиниан Бродманн опубликовал карты цитоархитектонических полей коры больших полушарий головного мозга. Бродман впервые создал карты коры. Впоследствии О. Фогт и Ц. Фогт (1919-1920 гг.) с учётом волоконного строения описали в коре головного мозга 150 миелоархитектонических участков. В Институте мозга АМН СССР И. Н. Филипповым и С. А. Саркисовым были созданы карты коры головного мозга, включающие 47 цитоархитектонических полей .
Несмотря на критику , поля Бродмана являются самыми известными и наиболее часто цитируемыми при описании нейрональной организации коры головного мозга и её функций.
Функционирование коры головного мозга, как и всего организма в целом, осуществляется в двух основных, четко отличающихся друг от друга состояниях: бодрствования и сна .
Реализация функций психических процессов, вовлекает весь мозг, и обеспечивается взаимосвязанной, скоординированной деятельностью многих участков коры головного мозга. Тем не менее, существуют области, в которых реализация конкретных функций в значительной степени локализуется. Поражение, недоразвитость, либо низкая активность таких участков вызывает нарушения выполнения локализованных в них функций. Так по функциям приёма сенсорных сигналов, формировании и управления движениями и обработки информации, кора головного мозга разделяется на три зоны: сенсорную, моторную и ассоциативную. При этом сенсорная зона обеспечивает приём и обработку сигналов органов чувств, моторная отвечает за произвольные движения. Функция ассоциативной зоны - связывать между собой активность сенсорных и моторных зон. Ассоциативная зона прогнозирует, получает и перерабатывает информацию из сенсорной зоны и, через моторную, инициирует и формирует целенаправленное осмысленное поведение.
Нейрохирург Уайлдер Пенфилд , занимавшийся хирургическим лечением эпилепсии , к середине XX века составил карту динамической локализации функций коры головного мозга. Во множество учебников вошла его схема, где изображена представимость различных органов человека в его головном мозгу .
В настоящее время основным инструментом для определения нейронных сетей, участков головного мозга, активизирующихся при определенных условиях, является позитронно-эмиссионная томография . В ходе таких исследований определяются нейронные сети , вовлекающиеся в реализацию конкретных функций. В частности, так была выявлена сеть пассивного режима работы мозга , активная в состоянии свободной, не целенаправленной умственной деятельностью, когда человек бездействует, отдыхает, грезит наяву или погружён в себя, а не связан с выполнением какой-либо задачи внешнего мира.
Сенсорная кора (зона) - это часть коры больших полушарий, которая получает и обрабатывает информацию от органов чувств . Области коры головного мозга, которые получают сенсорные сигналы от таламуса , называются первичными сенсорными зонами. Сигналы от органов зрения, слуха и осязания поступают в первичную зрительную , и кору, соответственно. Как правило, два полушария получают информацию от противоположной (контралатеральной) стороны тела . Например, правая первичная соматосенсорная кора получает информацию от левых конечностей, а правая зрительная кора получает информацию от рецептивного поля левого зрительного нерва. Топография сенсорных зон коры отражает топографию рецептивного поля соответствующего органа чувств и называется . Например, соседние точки в первичной зрительной коре соответствуют соседним точкам сетчатки . Эта карта называется . Таким же образом, существует в первичной слуховой коре и в первичной сенсорной коре. Последняя топографическая карта тела на была проиллюстрирована как искаженное изображение человека, соматосенсорного , где размер различных частей тела отражает относительную плотность их иннервации. Областям с большим количеством сенсорной иннервации, таким, как кончики пальцев и губы, отвечают большие области коры для обработки более тонких ощущений.
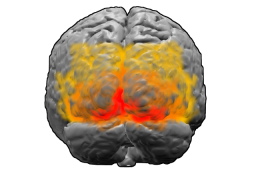
Действия вопреки страху , аффективное, эмоционально насыщенное поведение сопровождаются активацией так называемой подколенной области передней части поясной извилины головного мозга (subgenual anterior cingulate cortex — sgACC). Чем больше страх, тем сильнее активизируется эта область мозга. При этом одновременно подавляется активность височных долей головного мозга .
В июле 2016 на сайте журнала Nature была опубликована информация о карте коры головного мозга, составленной в результате исследований, проведенных Дэвидом Ван Эссеном (David Van Essen) и его коллегами из Медицинской школы Университета Дж. Вашингтона. Использование алгоритмов машинного обучения позволило идентифицировать 180 структурных участков коры головного мозга, вовлеченных в выполнение различных функций, занимающих 96.6% площади коры, включая 97 прежде неизвестных. В качестве исходных данных использовались изображения мультимодальной магнитно-резонансной томографии головного мозга 210 здоровых подопытных обоих полов, выполнявших простые задания, полученные в ходе реализации проекта по установлению полной «карты» структурных взаимосвязей мозга «Коннектом человека» (Human Connectome Project, HCP) .
Обнаружено, что центры конфликта находятся в передней поясной коре, неприятия в островковой доле .
Усвоение и понимание письменной и устной речи осуществляется частью коры головного мозга, размещённой в заднем отделе верхней височной извилины доминантного полушария, называемой областью Вернике .
Моторная организация речи, преимущественно связанная с фонологической и синтаксической кодификациями, обеспечивается работой центра Брока , участка коры, находящегося в задненижней части третьей лобной извилины левого полушария (у правшей).
Количественные значения активируют участки, расположенные в лобных и задних отделах теменных долей, головного мозга. Одним из ключевых мест является внутритеменная борозда, где представлен семантический смысл чисел . У людей, страдающих дискалькулией - неспособностью к изучению арифметики, данный участок мозга меньше, чем у здоровых людей, и недостаточно активен. В состав сетей, связанных с решением математических задач, входит часть центра Брока.
|
Лобная
доля |
|
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
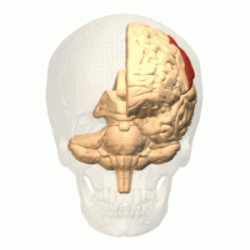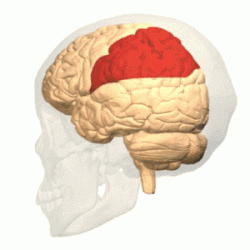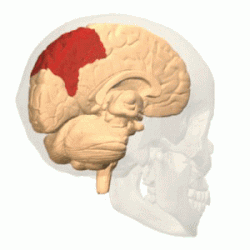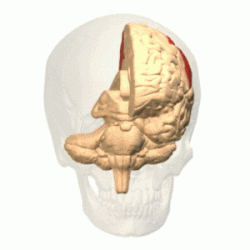
Теменная
доля |
|
||||||||||||||
|
Затылочная
доля |
|
||||||||||||||
|
Височная
доля |
|
||||||||||||||
|
Междолевые
борозды |
|
||||||||||||||
|
|||||||||||||||
|
Островковая
доля |
|
||||||||||||||
| Прочее | |||||||||||||||