Interested Article - Вентральная область покрышки

- 2020-01-15
- 1
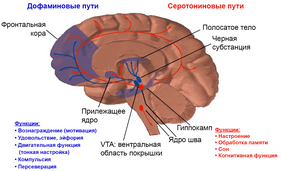
Вентральная область покрышки (ПОВ, англ. ventral tegmental area, VTA ) — часть среднего мозга , расположенная дорсомедиально по отношению к чёрному веществу и вентрально по отношению к красным ядрам . Является началом мезокортикального и мезолимбического дофаминовых путей. Вентральная область покрышки широко вовлечена в системы вознаграждения , а точнее сама по себе является скоплением множества нервных путей. Вентральная область покрышки также вовлечена в формирование зависимостей от таких веществ как никотин , кокаин , героин , амфетамин .
Структура
Нейробиологи часто сталкивались с большими трудностями при различении ВОП у людей и других приматов мозгов от чёрной субстанции и окружающих ядер. Первоначально вентральная область покрышки была обозначена как "ядро", но с течением времени "площадь" стала более подходящим термином из-за гетерогенных цитоархностических особенностей региона и отсутствия четких границ, отделяющих его от смежных областей. В связи с селективными лимбическими афферетами к ВТС, ячейкам ВТА дается обозначение А10, чтобы отличать их от окружающих клеток.
Местоположение
ВОП находится в середине мозга между несколькими другими основными областями, некоторые из которых описаны здесь. Сосцевидные тела и задний гипоталамус , включенные в диэнцефалон , расположены ростральнее (кпереди от) ВОП. Красное ядро расположено в боковом направлении, а глазодвигательные волокна расположены вентромедиально к ВОП. Варолиев мост и задний мозг находятся каудально(нижней части,ниже) от ВОП.Корешок расположен сбоку от ВОП.
Подразделения
В 1987 году Oades идентифицировал четыре первичных ядра в группе клеток VTA A10: nucleus paranigralis (Npn), nucleus parabrachialis pigmentosus (Npbp), nucleus interfascicularis (Nif) и nucleus linearis (Nln) caudalis и rostralis. В настоящее время ученые разделяют ВОП на четыре аналогичные зоны, которые называются паранигральным ядром (ПН), парабрахиальной пигментной областью (ПБП), парафасциальной ретрофлексной областью (ПФР) и ростромедиальным тегментальным ядром (РМТг), которые примерно соответствуют предыдущим подразделениям. Некоторые определения ВОП также включают ядра средней линии (то есть межфасциальное ядро, ростральное линейное ядро и центральное линейное ядро).
ПН и ПБП богаты дофаминергическими клетками, в то время как другие две области имеют низкую плотность этих нейронов. ПФР и РМТг содержат низкую плотность тирозингидроксилазы (ТГ)-положительных клеточных телец, которые малы по размеру и слабо окрашены; РМТг состоит в основном из гамкаэргических клеток. С другой стороны, ПН и ПБП состоят в основном из средних и крупных Т-позитивных клеточных телец, которые окрашиваются умеренно.
Входы
Почти все области, получающие проекции от проекта ВОП, возвращаются к нему. Таким образом, вентральная тегментальная область взаимно связана с широким спектром структур по всему мозгу , предполагая, что она играет определенную роль в контроле функций в филогенетически более новым и высокоразвитым неокортексе, а также в филогенетически более старых лимбических областях.
ВОП представляет собой гетерогенную область, состоящую из множества нейронов, которые характеризуются различными нейрохимическими и нейрофизиологическими свойствами. Таким образом, глутаматергический и ГАМКергический входы не являются исключительно ингибирующими или исключительно возбуждающими. ВОП получает глутаматергические афференты из префронтальной коры, педункулопонтинного тегментального ядра (ППТг), латеродорсального тегментального ядра, субталамического ядра, ядра ложа stria terminalis, верхнего колликулуса, периакведуктального серого, латерального хабенула, дорсального ядра рафа , латеральной гипоталамической и преоптической областей. Эти глутаматергические афференты играют ключевую роль в регуляции активации клеток ВОП. Когда глутаматергические нейроны активируются, скорость возбуждения дофаминовых нейронов увеличивается в ВОП и вызывает взрывное возбуждение. Исследования показали, что эти глутаматергические действия в ВОП имеют решающее значение для последствий злоупотребления наркотиками. Напротив, хвост вентральной тегментальной области (РМТг) проецируется на ВОП с ГАМКергическими афферентами, функционирующими как "главный тормоз" для дофаминовых путей ВОП .
ГАМКергические входы в ВОП также включают прилежащее ядро, вентральный паллидум, дорсальное ядро рафе , латеральный гипоталамус, периакведуктальный серый, ядро ложа терминальной стрии ростромедиальное тегментальное ядро (РМТг). Латеральная хабенула также может оказывать ингибирующее действие на дофаминергические нейроны в ВОП путем возбуждения РМТг гамкаэргических нейронов, который, как полагают, играет важную роль в вознаграждении ошибки прогнозирования . Субпаллидные афференты в ВОП в основном являются гамкаэргическими и, таким образом, ингибиторными. Существует существенный путь от субпаллидной области к ВОП. Когда этот путь расторможен, увеличение высвобождения дофамина в мезолимбическом пути усиливает локомоторную активность.
Существуют также холинергические входы в ВОП, хотя и менее изученные, чем глутаматергические и ГАМКергические входы. Оптогенетические исследования на мышах, изучающих холинергические входы из тегментального ядра педункулопонтина (ППТг) и латеродорсального тегментального ядра, показывают, что эти цепи усиливают разрядные свойства нейронов ВОП, предполагая модулирующее влияние на схемы вознаграждения .
Выходы
Двумя первичными проекциями эфферентных волокон ВОП являются мезокортикальный и мезолимбический пути, которые соответствуют префронтальной коре и прилежащему ядру соответственно. Кроме того, эксперименты на грызунах выявили мезогабенулярный путь, состоящий из нейронов ВОП, которые выделяют не дофамин, а глутамат и ГАМК . Другие проекции ВОП, которые используют дофамин в качестве основного нейромедиатора, перечислены ниже .
-
Проекции вентральной тегментальной области (VTA)
- ВОП → Миндалевидное тело
- ВОП → Энторинальная кора
- ВОП → Поясная извилина
- ВОП → Гиппокамп
- ВОП → Прилежащее ядро
- ВОП → Обонятельная луковица
- ВОП → Префронтальная кора
Развитие
Поскольку они развиваются из общей эмбриональной ткани и частично накладываются друг на друга в своих проекционных полях, дофаминергические группы клеток не имеют четких анатомических границ. Во время развития мозга млекопитающих нейроны черной субстанции (SN) и ВТА первоначально проецируются в дорсолатеральный и вентромедиальный стриатум. Однако при рождении дофаминергические нейроны SN проецируются исключительно в дорсолатеральный стриатум, а дофаминергические нейроны VTA проецируются исключительно в вентромедиальный стриатум. Это сокращение связей происходит через устранение ненужных коллатералей.
Функции
Как было сказано выше, ВОП, в частности дофаминовые нейроны ВОП, выполняют несколько функций в системе вознаграждения, мотивации, познании, наркомании могут быть в центре внимания некоторых психических расстройств. Было также показано, что ВОП обрабатывает различные типы эмоций, исходящих из миндалевидного тела, где он также может играть роль в избегании и обусловливании страха . Электрофизиологические записи показали, что нейроны ВПО реагируют на новые стимулы, неожиданные награды и сенсорные сигналы, предсказывающие вознаграждение. Схема срабатывания этих ячеек согласуется с кодированием ошибки ожидания вознаграждения.
В 2006 году МРТ-исследования Хелен Фишер и ее исследовательская группа обнаружила и задокументировала различные эмоциональные состояния, связанные с интенсивной любовью, коррелирующие с активностью в ВОП, что может помочь объяснить навязчивое поведение отвергнутых партнеров, поскольку это разделяется системой вознаграждения. Поведение совместного использования гнезд связано с повышенной экспрессией V1aR в ВОП недавно спаренных зебровых вьюрков. Однако экспрессия V1aR не была связана с частотой женских направленных песен, что может указывать на избирательную роль вазотоцина в ВОП в поддержании парного поведения по сравнению с поведением ухаживания.
Наличие щелевых соединений
Было показано, что ВОП имеет большую сеть ГАМКергических нейронов, которые связаны между собой через щелевые соединения. Эта сеть обеспечивает электрическую проводимость, которая значительно быстрее, чем химическая проводимость сигналов между синапсами, хотя и менее пространственно точна .
Примечания
- // SpringerReference. — Berlin/Heidelberg: Springer-Verlag.
- Zdravko Vitošević, Slobodan Marinković, Mila Ćetković, Bojan Štimec, Vera Todorović. // Anatomical Science International. — 2012-12-15. — Т. 88 , вып. 2 . — С. 70–82 . — ISSN . — doi : .
- Volker Arnd Coenen, Lena Valerie Schumacher, Christoph Kaller, Thomas Eduard Schlaepfer, Peter Christoph Reinacher. // NeuroImage: Clinical. — 2018. — Т. 18 . — С. 770–783 . — ISSN . — doi : .
- Romain Bourdy, Michel Barrot. (англ.) // Trends in Neurosciences. — 2012-11. — Vol. 35 , iss. 11 . — P. 681–690 . — doi : . 8 марта 2021 года.
- M. Barrot, S. R. Sesack, F. Georges, M. Pistis, S. Hong. (англ.) // Journal of Neuroscience. — 2012-10-10. — Vol. 32 , iss. 41 . — P. 14094–14101 . — ISSN . — doi : .
- Mitsuko Watabe-Uchida, Neir Eshel, Naoshige Uchida. (англ.) // Annual Review of Neuroscience. — 2017-07-25. — Vol. 40 , iss. 1 . — P. 373–394 . — ISSN . — doi : . 11 июля 2021 года.
- Daniel Dautan, Albert S Souza, Icnelia Huerta-Ocampo, Miguel Valencia, Maxime Assous. (англ.) // Nature Neuroscience. — 2016-08. — Vol. 19 , iss. 8 . — P. 1025–1033 . — ISSN . — doi : . 1 ноября 2021 года.
- ↑ Eric J. Nestler. . — 2nd ed. — New York: McGraw-Hill Medical, 2009. — 1 online resource с. — ISBN 978-0-07-164119-7 , 0-07-164119-X, 0-07-148127-3, 978-0-07-148127-4, 978-1-281-79174-0, 1-281-79174-1.
- Mihai Nechifor. // Magnesium Research. — 2008-03. — Т. 21 , вып. 1 . — С. 5–15 . — ISSN . 16 апреля 2021 года.
- Alice M. Stamatakis, Joshua H. Jennings, Randall L. Ung, Grace A. Blair, Richard J. Weinberg. (англ.) // Neuron. — 2013-11. — Vol. 80 , iss. 4 . — P. 1039–1053 . — doi : . 25 мая 2021 года.
- David H Root, Carlos A Mejias-Aponte, Shiliang Zhang, Hui-Ling Wang, Alexander F Hoffman. (англ.) // Nature Neuroscience. — 2014-11. — Vol. 17 , iss. 11 . — P. 1543–1551 . — ISSN . — doi : . 12 августа 2021 года.
- S. Mingote, N. Chuhma, S. V. Kusnoor, B. Field, A. Y. Deutch. (англ.) // Journal of Neuroscience. — 2015-12-09. — Vol. 35 , iss. 49 . — P. 16259–16271 . — ISSN . — doi : .
- ↑ Wei Tang, Olexiy Kochubey, Michael Kintscher, Ralf Schneggenburger. (англ.) // The Journal of Neuroscience. — 2020-05-13. — Vol. 40 , iss. 20 . — P. 3969–3980 . — ISSN . — doi : .
- J. P. Fadok, T. M. K. Dickerson, R. D. Palmiter. (англ.) // Journal of Neuroscience. — 2009-09-09. — Vol. 29 , iss. 36 . — P. 11089–11097 . — ISSN . — doi : .
- David W. Allison, Allison J. Ohran, Sarah H. Stobbs, Manuel Mameli, C. Fernando Valenzuela. (англ.) // Synapse. — 2006-07. — Vol. 60 , iss. 1 . — P. 20–31 . — ISSN . — doi : . 9 июля 2021 года.
|
|
В статье
не хватает
ссылок на источники
(см.
рекомендации по поиску
).
|
- 2020-01-15
- 1