Маас
- 1 year ago
- 0
- 0

Мозазавры ( лат. Mosasauridae ) — семейство вымерших морских ящеров (Lacertilia) из надсемейства ( или ) . Довольно близкие родственники современных варанов .
Первые ископаемые остатки мозазавров были найдены в известняковом карьере в Маастрихте на Маасе в 1764 году. В настоящее время ископаемые остатки представителей этого семейства уже были обнаружены на всех континентах, включая Антарктиду .
Мозазавры представляли собой большую группу верхнемеловых морских рептилий, главным образом хищников крупных или средних размеров. Большинство известных видов населяло тёплые, мелководные моря, широко распространённые в позднем меловом периоде. В силу своей специализации, мозазавры очень сильно отличались от современных ящериц: строение их внутренних органов больше напоминало таковое у китообразных, они были живородящими , имели высокие темпы обмена веществ и были теплокровными.
Мозазавры, вероятно, произошли от вымершего семейства водных ящериц ( ). В течение последних 20 млн лет мелового периода ( туронский — маастрихтский века) мозазавры вытеснили всех своих конкурентов в лице крупных ламнообразных акул и последних плиозавров , и стали доминирующими морскими хищниками своего времени. Они исчезли вместе с динозаврами и птерозаврами в результате массового вымирания в конце мелового периода, произошедшего 66 млн лет назад.

Первое официальное открытие частичного ископаемого черепа мозазавра произошло в 1764 году, и было сделано работниками в известняковом карьере на горе , недалеко от голландского города Маастрихта . Тем не менее, эта находка осталась малоизвестной и только второе обнаружение фрагментарного черепа мозазавра, произошедшее между 1770 и 1774 годами, привлекло к себе большое количество внимания. К. Гофман, хирург и коллекционер ископаемых, обсуждал череп мозазавра вместе с самыми влиятельными учёными своего времени, делая находку всё более и более известной. Первоначальным владельцем черепа, тем не менее, был каноник собора в Маастрихте.
Когда французские революционные войска заняли Маастрихт в 1794 году, тщательно спрятанная окаменелость была найдена и перевезена в Париж. После этого мозазавр был описан различными учёными как рыба, крокодил или кашалот. Первым, кто понял, что на самом деле это была гигантская морская ящерица, был голландский ученый П. Кампер в 1799 году. В 1808 году Жорж Кювье подтвердил этот вывод. Название «мозазавр» («ящер с реки Маас ») животное получило в 1822 году, а видовое название Mosasaurus hoffmannii — в 1829.

Известняковый карьер Маастрихта стал настолько известным за счёт открытия мозазавра, что в честь города был назван ярус , включающий в себя последние 6 миллионов лет мелового периода.
В 2014 году команда палеонтологов обнаружила фрагмент черепа крупного мозазавра в Италии. Команда оценила, что череп этого мозазавра достигал приблизительно 1,45 м в длину, что соответствует длине более 14,5 метров и делает этого мозазавра самой большой рептилией, когда-либо известной из Италии. Поскольку время его существования датируется кампанским ярусом, это один из самых ранних представителей рода Mosasaurus . Возможно, этот мозазавр образует сестринскую группу с Mosasaurus hoffmannii, отличаясь от своего более позднего родственника, в первую очередь, строением зубов .
В 2015 году учёные обнаружили окаменелости нового вида рода в Японии, теперь известного как Phosphorosaurus ponpetelegans . Скорее всего, этот мозазавр жил в глубоком океане, где охотился на рыбу и кальмаров, учитывая его большие глазницы и более развитое бинокулярное зрение . В 2023 г. в Мексике обнаружен ранее неизвестный вид мозазавра, названный Yaguarasaurus regiomontanus
Уровень моря был относительно высоким на протяжении позднего мелового периода, в результате чего ископаемые остатки морских животных того времени зачастую могут быть найдены в местах, ныне являющихся сушей. Окаменелости мозазавров были найдены в Нидерландах , Бельгии , Дании , Португалии , Швеции , Испании , Франции , Германии , Польши , Чехии , Болгарии , Великобритании , США , Канаде , России ( Поволжье ) , Украине , Казахстане , Азербайджане , Японии , Египте , Израиле , Иордании , Сирии , Турции , Мексике , Колумбии , Бразилии , Перу , Чили , Нигере , Анголе , Марокко , Австралии , Новой Зеландии и на острове Вега у берегов Антарктиды . Зуб мозазавра, классифицируемого как , известен с острова Тимор . Однако, филогенетическое размещение этого вида является неопределённым, и на самом деле он может даже и не быть мозазавром .

Животное, предположительно близкое к общему предку мозазавров, было найдено в отложениях нижнего мела из Японии . Это была мелкая водная ящерица ( Kaganaias ), обладавшая телом змеевидной формы и короткими конечностями.
На основании некоторых признаков, таких как дополнительный ряд зубов на птеригоидах, слабо сочленённая нижняя челюсть, видоизменённые конечности и предполагаемые ранее методы локомоции , многие исследователи прошлого считали, что современные змеи имели общего морского предка с мозазаврами. Так, в 1869 году Эдвард Дринкер Коуп ввёл термин , для того, чтобы их объединить . Эта теория держалась более столетия, пока некоторые находки, такие как роющая змея из Южной Америки, названная Najash rionegrina , не поставили под сомнение гипотезу морского происхождения змей и их близкого родства с мозазаврами.
Скелет описанного в 2005 году примитивного мозазавра Dallasaurus turneri имеет ряд признаков, присутствующих в скелетах развитых мозазавров подсемейства Mosasaurinae и в скелетах их предполагаемых предков, таких как . Но при этом даллазавр сохраняет конечности, похожие по строению и функционалу на конечности айгиалозавра и современных сухопутных чешуйчатых, в то время как у его потомков — мозазавров — конечности преобразовывались в веслообразные плавники .
Филогенетический анализ 2005 года указал на то, что мозазавры, имеющие видоизменённые в плавники конечности, не образуют клады, и не включают в себя такие таксоны, как Dallasaurus, , , , и . Видоизменение конечностей произошло независимо в трёх разных группах мозазавров (Halisaurinae, Mosasaurinae и группы, состоящей из подсемейств Tylosaurinae и Plioplatecarpinae) . В дальнейшем исследования 2007 и 2012 года подтвердили эту гипотезу , но авторы одного из исследований 2012 года пришли к выводу о том, что независимое видоизменение конечностей в плавники произошло только у двух разных групп мозазавров (у Mosasaurinae и у клады, содержащей Halisaurinae, Tylosaurinae и Plioplatecarpinae) . С другой стороны, в одной из работ 2011 года и вовсе была возвращена идея о том, что видоизменение конечностей мозазавров произошло в рамках одной группы, от которой берут своё начало все остальные . Если изначальная гипотеза 2005 года верна, то семейство Mosasauridae является полифилетической группой и должно быть расформировано, либо в него должны быть включены таксоны, не имеющие видоизмененные в плавники конечности .
Точное филогенетическое положение клады, содержащей мозазавров и их ближайших родственников в отряде чешуйчатых, на данный момент остаётся неопределённым. Старые кладистические анализы указывали их как тесно связанную со змеями группу , но на данный момент это было опровергнуто . Морфологический анализ, проведённый в 2008 году, указал на то, что мозазары являются варанообразными ящерицами, тесно связанными с современными варанами. Анализ 2011 года, основанный на морфологических особенностях групп Mosasauridae, Aigialosauridae и , указал варанов и ядозубов в качестве ближайших родственников мозазавров; но анализ, основанный на сочетании морфологических и молекулярных данных, с другой стороны, указал варанов и безухих варанов более близкими родственниками мозазавров, чем ядозубов и крокодилового шинизавра . Морфологический анализ, проведённый 2012 году, внезапно указал мозазавров и их родственников как базальных членов клады (содержащей все таксоны, более тесно связанные с Gecko gecko и Scincus scincus , чем с Iguana iguana ), которые не принадлежат к кладе . Филогенетическое положение мозазавров по методике исследования 2012 года, как выяснилось в дальнейшем, сильно зависит от того, какие таксоны будут включены или исключены из этого анализа. Когда группа Mosasauridae была исключена из анализа, Aigialosauridae и Dolichosauridae были восстановлены в кладу Scleroglossa, образуя сестринскую группу по отношению к кладе, содержащей змей, двуходок , червеобразных ящериц и безногих ящериц . Когда Mosasauridae были возвращены в анализ, а чешуйчатые с отсутствующими конечностями (кроме змей) были исключены, Mosasauridae, Aigialosauridae и Dolichosauridae вошли в кладу Scleroglossa, формируя родственную группу со змеями . Более детальный анализ, проведённый в том же самом году, указал что положение клады, содержащей мозазавров и их ближайших родственников, внутри отряда чешуйчатых является нестабильным и по-разному восстанавливается вне Scleroglossa или рядом с безногими формами .

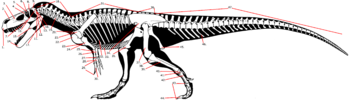
Мозазавры имели форму тела, напоминающую таковую у современных варанов, но более вытянутую и обтекаемую. Головы многих мозазавров обычно были относительно крупнее по отношению к телу, чем у варанов. Кости конечностей были сокращены, а их длинные пальцы формировали ласты, помогавшие животным поворачивать при движении. Хвосты были расширены и уплощены.
До недавнего времени считалось, что мозазавры использовали относительно примитивный метод плаванья, свойственный засадным охотникам, таким как мурены , крокодилы и морские змеи : отталкивались от воды, извиваясь всем телом. Однако, современные исследования показали, что развитые мозазавры имели гипоцеркальные хвостовые плавники , напоминающие таковые у , ихтиозавров и некоторых акул. Вместо того, чтобы активно двигаться, передняя часть тела мозазавров оставалась сравнительно неподвижной, чтобы уменьшить лобовое сопротивление воды, и все двигательные усилия приходились на хвост. Таким образом, эти животные не были специализированными засадными охотниками и могли активно преследовать свою жертву в толще воды .

Ранние реконструкции некоторых мозазавров ( тилозавров ) изображали их с длинными гребнями вдоль всего тела, за которые ошибочно были приняты хрящи от трахеи. Когда эта ошибка была обнаружена и устранена, изображаемые с подобного рода наростами мозазавры уже получили достаточно широкое распространение .
Самый маленький известный представитель семейства — даллазавр Dallasaurus turneri , был менее 1 метра в длину. Однако, крупные мозазавры были более распространены — большинство видов обычно вырастало свыше 6 метров в длину. Mosasaurus hoffmannii , крупнейший из известных науке мозазавров, мог вырастать более 17 метров в длину . В настоящее время, самый большой публично выставленный скелет мозазавра находится на палеонтологической выставке в Мордене, Манитоба, Канада. Это образец по прозвищу «Брюс», длинной около 13 метров .
Костная структура развитых мозазавров демонстрирует хорошую адаптацию к активному плаванью, в то время как примитивные мозазавры, такие как даллазавр, имеют костную структуру, напоминающую таковую у наземных или полуводных ящериц . Мозазавры обладали , как крокодилы. Период смены зубов у платекарпа (образец JAG 0006 t) составлял около 260, у прогнатодона (образец JAG 0005 t) — 374 дня, а у тилозавра (образец JAG 0007 t) — около 593 дней, в то время как у современного им крокодиломорфа (образец JAG 0001 t) — около 240 дней .

Несмотря на многочисленные ископаемые остатки различных мозазавров, происходящих со всего мира, о их кожном покрове известно немного. Несколько образцов, тем не менее, сохраняют окаменевшие отпечатки чешуи . До открытия хорошо сохранившихся отпечатков из позднего маастрихта Иордании , кожный покров мозазавров описывался в основном на базе ископаемых остатков довольно примитивных мозазавров из сантона-нижнего кампана, таких как знаменитый образец тилозавра KUVP—1075 из штата Канзас . Материал из Иордании показывает, что тела мозазавров, а также перепонки между пальцами на передних и задних конечностях, были покрыты мелкими, перекрывающимися, ромбовидной формы чешуйками, напоминающие таковые у некоторых змей. Также как и у многих современных рептилий, чешуя мозазавров была разнообразной по типу и размеру на различных участках тела. У обнаруженных образцов имелось два основных типа чешуй: килеватая чешуя на верхних участках тела и гладкая чешуя на нижних .

Сравнительно недавно был обнаружен окаменелый образец Platecarpus tympaniticus, сохранивший не только отпечатки кожи, но и внутренних органов. Несколько красноватых участков в ископаемом образце могут представлять собой сердце , легкие и почки . Трахея также сохранилась, вместе с частью того, что может быть сетчаткой глаза. Размещение почек дальше вперед в области живота больше напоминает таковое у китообразных , нежели у варанов. Как и у китообразных, бронхи, ведущие к легким, у мозазавров шли параллельно друг—другу, а не перекрывались, как у варанов и других сухопутных и полуводных ящериц. Эти анатомические признаки скорее всего являются следствием полной приспособленности к морскому образу жизни .

В 2011 году белок коллаген был извлечен из плечевой кости прогнатодона .
Окрас чешуи мозазавров был неизвестен до 2014 года, пока не были обнаружены меланины в окаменевшей чешуе мозазаврида вида Tylosaurus nepaeolicus . Исследования показали, что он скорее всего имел темную дорсальную поверхность тела и более светлую вентральную , примерно как современная белая акула или кожистая черепаха . Причем, для вымерших предков последней окрас был определён аналогичным образом .
Исследования, опубликованные в 2016 году, свидетельствуют о том, что мозазавры были теплокровными животными. Сравнив изотопный состав зубов трёх родов мозазавров с данными, известными для заведомо холоднокровных рыб рода и безусловно теплокровных птиц рода Ichthyornis , американские учёные получили следующую температуру тела для представителей трех родов мозазавров: 33,1 °C у Clidastes , 34,3 °C у Tylosaurus , 36,3 °C у Platecarpus . Температура энходуса при этом составляла 28,3 °C, ихтиорниса — 38,6 °C . Более поздние и развитые мозазавры, такие как Mosasaurus и Plotosaurus , возможно, имели ещё более высокий обмен веществ.
Исследования изотопного состава зубов 2010 года от А. Бернарда и соавторов, указывали диапазон поддерживаемой температуры тела мозазавров в (35±2 — 39±2) °С, но не исключали того, что она могла несколько зависеть от температуры окружающей среды . Микроструктура костной ткани мозазавров демонстрирует промежуточный уровень метаболизма между современной частично теплокровной кожистой черепахой и ихтиозаврами или плезиозаврами, теплокровность и очень высокий уровень метаболизма которых считаются доказанными .
Онтогенез мозазавров известен довольно плохо, во многом потому что ископаемые остатки детенышей мозазавров редки и при находке их раньше часто принимали за гесперорнисообразных птиц. Однако, исследование некоторых образцов мозазавров, обнаруженных более века назад, свидетельствуют о том, что мозазавры рожали живых детенышей, и что по крайней мере в некоторых случаях детеныши проводили свои первые годы жизни в открытом океане, а не в защищенных яслях или мелководных областях. Забота о потомстве для мозазавров кажется крайне маловероятной, учитывая их активный образ жизни и отсутствия подобного поведения у современных варанов . В процессе роста детеныши, вероятно, занимали разные экологические ниши, и тем самым избегали конкуренции со взрослыми особями.
Из-за высокого уровня обмена веществ мозазавры росли быстрее, чем современные вараны . Несмотря на крупный размер, для того, чтобы достичь половой зрелости, клидастам , тилозаврам и платекарпам, по оценкам, требовалось в районе 5—7 лет: столько же, сколько и современному комодскому варану .

Мозазавры заняли многие экологические ниши в морях мелового периода. Главным образом ниши активных хищников крупных и средних размеров. Гигантские мозазавры, адаптированные к убийству крупной добычи, располагались на вершине пищевой пирамиды позднемеловых морей.
Из-за быстрого метаболизма, мозазаврам требовалось относительно большее количество пищи, чем современным рептилиям. Ранние мозазавры имели раздвоенные нижние челюсти и множество подвижных сочленений в черепе, отдаленно напоминая в этом плане змей. Такой краниальный кинетизм позволял им проглатывать относительно крупные пищевые объекты целиком, а зубы на птеригоидах, как у некоторых мурен , помогали мозазаврам проталкивать в глотку большие куски пищи. Однако, кинетизм черепа некоторых более поздних форм был сильно снижен и зубы многих из них приобретали режущий морфотип, что позволяло развитым мозазавром с легкостью выпиливать куски мяса из очень крупной добычи, а также выдерживать нагрузки, возникающие при её сопротивлении. Но некоторые мозазавры, такие как глобиденс , развили довольно необычные адаптации для кормления. Зубы этого мозазавра были шаровидной, удобной для раскусывания раковин моллюсков формы, подобно зубам триасовых плакодонтов .
Известно содержимое желудка некоторых мозазавров: один образец тилозавра ( Tylosaurus proriger ) содержал в области желудка ископаемые остатки нелетающей морской птицы гесперорниса ( Hesperornis ), крупных костных рыб, меньшего мозазавра платекарпа ( Platecarpus tympaniticus ) и акулы. В желудках других тилозавров ( T. proriger и T. nepaeolicus ) были найдены ископаемые остатки огромной акулы — гибодонта Ptychodus mortoni , клидаста ( Clidastes sp.), ещё одного платекарпа ( Platecarpus planifrons ), крупного ксифактина ( Xiphactinus audax ) и короткошеего плезиозавра рода . В желудке мелкого рода мозазавра ( Mosasaurus missouriensis, около 6,5 м длиной) были найдены ископаемые остатки метровой костной рыбы, перекушенной в нескольких местах , в то время как желудок другого мозазавра этого рода содержал ископаемые остатки неидентифицированного мозазавра из подсемейства и плохо сохранившееся животное, не являющееся мозазавром. В области желудка у гайнозавров были найдены ископаемые остатки плиоплатекарпуса ( Plioplatecarpus sp.), неидентифицированного мозазавра и кости морской черепахи . Последние, вероятно, были проглочены гайнозавром целиком или выедены из панциря, поскольку его гибкие челюсти и лезвиеподобные зубы хорошо подходят для разрезания шкуры и мяса, но не для прокусывания твердых защитных покровов. Содержимое желудка некрупного прогнатодона ( Prognathodon overtoni , около 5.9 м длиной) включало в себя очень крупных костных рыб, свыше 25 % от общей длины тела мозазавра, черепахи с панцирем длиной около 60 см и, возможно, головоногого моллюска . В области желудка Prognathodon kianda , в то же время, были найдены ископаемые остатки трех других мозазавров, включая молодую особь его вида . Мелкие мозазавры с очень подвижными черепами — платекарпы, содержали в области желудка только костных рыб длиной до 1,2 м (хотя строение их зубов предполагает возможность порой питаться и другими, более крупными животными). В области желудка плотозавра — сравнительно крупного, но специализированного под нишу ихтиофага вида, также содержалась только относительно мелкая рыба . Между зубами мозазавра из рода были найдены застрявшие куски морского ежа. Желудок плиоплетекарпуса ( Plioplatecarpus sp.) содержал в себе двух белемнитов .
Некоторые виды жертв мозазавров известны по конкретным отпечаткам на них, а не по содержимому желудков хищников. Так, необычные отверстия были найдены на раковинах аммонитов (в основном и ) . Когда-то они идентифицировались как повреждения от паразитов, но позже было установлено, что треугольная форма отверстий на некоторых из них, их размер и наличие с обеих сторон, соответствует верхней и нижней челюсти при укусе среднего или небольшого мозазавра, скорее всего платекарпа. Неизвестно, поедали ли аммонитов более крупные мозазавры, хотя отсутствие доказательств тому даже в таких местах, откуда известно большое количество ископаемых остатков аммонитов и крупных мозазавров, говорит о том, что головоногие моллюски скорее всего не были энергетически-выгодной пищей для более крупных мозазавров . Панцирь гигантской морской черепахи ( Allopleuron hofmanni ) имеет на себе следы от зубов крупного мозазавра рода Mosasaurus или Prognathodon . Следы от зубов мозазавров также были найдены на костях некоторых крупных длинношеих плезиозавров и известен плавник молодого короткошеего плезиозавра, буквально разрезанный зубами небольшого прогнатодона . Из Новой Зеландии сообщается обнаружение мозазавра, сцепившегося с плезиозавром Mauisaurus haasti . Была описана мантия гигантского головоногого моллюска, носящая на себе разрезы от зубов неопознанного мозазавра . Кости гадрозавра из Аляски носят на себе следы поедания тилозавром, что, скорее всего является результатом употребления в виде падали вынесенного в море животного.
Многие кости мозазавров, даже больших представителей наиболее крупных видов, носят на себе следы от зубов других мозазавров. Известны находки как со следами заживления, так и без них. Это является прямым свидетельством агрессивного взаимодействия мозазавров друг с другом, выраженного в территориальных боях, каннибализме и случающихся межвидовых конфликтах, возможно, демонстрирующих хищничество . Большой интерес представляет находка 110—120 см черепа молодого мозазавра вида Mosasaurus hoffmanni, который скорее всего был пробит рострумом гайнозавра .
На костях мозазавров часто встречаются следы от зубов акул (в основном скваликораксов ), демонстрирующие поедание акулами мертвых тел гигантских морских ящеров . Однако, известно две находки следов от зубов кретоксирин на костях ранних мозазавров (представителей родов Clidastes , Tylosaurus или Platecarpus ), происходящих в основном из кампанского яруса Канзаса , которые носят на себе следы заживления, и тем самым предполагают то, что акула кусала живого мозазавра. Неизвестно, при каких обстоятельствах были получены данные ранения и какая сторона в действительности была инициатором . Известен образец ксифактина , в челюстях которого застрял детеныш мозазавра неопределенного вида .



Кладограмма мозазавров от Simões et al. (2017) :
| Mosasauridae |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|---|---|
| Таксономия |
|