Слепое разделение сигнала
- 1 year ago
- 0
- 0

Передача сигнала (сигнальная трансдукция, трансдукция, сигналинг, сигнализация, англ. signal transduction ) — в молекулярной биологии термин «Передача сигнала» относится к любому процессу, при помощи которого клетка превращает один тип сигнала или стимула в другой.
Существование сложных многоклеточных организмов возможно благодаря координации биохимических процессов, протекающих в их клетках. Основой такой координации служат и передача сигнала внутри отдельных клеток. Вместе это даёт возможность одной клетке контролировать поведение остальных.
В большинстве случаев передача сигнала внутри клетки представляет собой цепь последовательных биохимических реакций, осуществляемых ферментами , часть из которых активируется вторичными посредниками . Такие процессы обычно являются быстрыми: их продолжительность — порядка миллисекунд в случае ионных каналов и минут — в случае активации протеинкиназ и . Однако в некоторых случаях от получения клеткой сигнала до ответа на него могут проходить часы и даже сутки (в случае экспрессии генов ). Пути передачи сигнала, или сигнальные пути , часто бывают организованы как сигнальные каскады ( англ. signal cascade ): количество молекул белков и других веществ, принимающих участие в передаче сигнала, возрастает на каждом последующем этапе по мере удаления от первоначального стимула. Таким образом, даже относительно слабый стимул может вызывать значительный ответ. Это явление называется амплификацией сигнала . Оригинальный термин англ. signal transduction впервые появился в реферируемых журналах в 1974 году, а в названии статьи фигурировал в 1979 году.
Нарушения в системе передачи сигналов могут привести к развитию рака , аутоиммунных заболеваний и диабета . Понимание механизмов передачи сигнала внутри клетки может привести к разработке методов лечения этих заболеваний и даже созданию искусственных тканей .
Первичные посредники — это химические соединения или физические факторы ( квант света, звук, внешняя температура, механические воздействия - давление, вибрация, сдвиг и другие), способные активировать механизм передачи сигнала в клетке. По отношению к воспринимающей клетке первичные посредники являются экстраклеточными сигналами. Стоит отметить, что в качестве экстраклеточных стимулов могут выступать и молекулы, в изобилии присутствующие внутри клетки, но находящиеся в норме в очень низкой концентрации в межклеточном пространстве (например, АТФ или глутамат ). В зависимости от функций первичные посредники могут быть разделены на несколько групп:
Получение клеткой сигнала от первичных посредников обеспечивается особыми белками- рецепторами , для которых первичные посредники являются лигандами . Для обеспечения рецепторной функции молекулы белков должны отвечать ряду требований:
Клеточные рецепторы делятся на следующие классы:
Мембранные рецепторы распознают крупные (например, инсулин) или гидрофильные (например, адреналин) сигнальные молекулы, которые не могут самостоятельно проникать в клетку. Небольшие гидрофобные сигнальные молекулы (например, трийодтиронин , стероидные гормоны , CO, NO) способны проникать в клетку за счёт диффузии . Рецепторы таких гормонов обычно являются растворимыми цитоплазматическими или ядерными белками. После связывания лиганда с рецептором информация об этом событии передаётся дальше по цепи и приводит к формированию первичного и вторичного клеточного ответа .
Если внешняя сигнальная молекула воздействует на рецепторы клеточной мембраны и активирует их, то последние передают полученную информацию на систему белковых компонентов мембраны, называемую каскадом передачи сигнала . Мембранные белки каскада передачи сигнала подразделяют на:
Так действуют рецепторы, сопряженные с G-белками . Другие рецепторы (ионные каналы, рецепторы с протеинкиназной активностью) сами служат умножителями.
Вторичные посредники ( англ. second messenger ) — это низкомолекулярные вещества, которые образуются или высвобождаются в результате ферментативной активности одного из компонентов цепи передачи сигнала и способствуют его дальнейшей передаче и амплификации . Вторичные посредники характеризуются следующими свойствами: имеют небольшую молекулярную массу и с высокой скоростью диффундируют в цитоплазме ; быстро расщепляются и быстро удаляются из цитоплазмы. Ко вторичным посредникам относятся:
Иногда в клетке образуются и третичные посредники . Так, обычно ионы Ca 2+ выступают в роли вторичного посредника, но при передаче сигнала с помощью инозитолтрифосфата (вторичный посредник) выделяющиеся при его участии из ЭПР ионы Ca 2+ служат третичным посредником.
Передача сигнала предполагает примерно следующую схему:

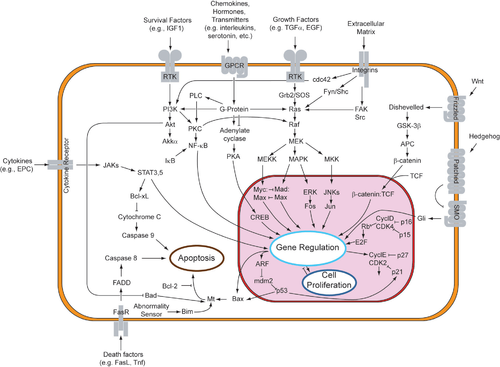
Иногда активация рецептора внешним стимулом сразу приводит к ответу клетки. Например, когда нейротрансмиттер ГАМК активирует свой рецептор, входящий в состав ионного канала на поверхности нейрона, канал начинает пропускать ионы хлорида, что приводит к изменению мембранного потенциала всей клетки. В других случаях активация рецептора лишь инициирует цепь событий, передающих регуляторный стимул внутри клетки через более или менее длинную цепь посредников. Такая цепь называется сигнальным путём .
относительно короток. Активация этого рецептора позволяет протеазе расщепить его, часть белка может затем попасть в клеточное ядро и выполнять функции регулятора транскрипции . В результате клетка начинает синтезировать другие белки и изменяет своё поведение. Более сложный сигнальный путь инициируют факторы роста. Некоторые из их рецепторов являются киназами и после активации фосфорилируют себя и другие белки, тем самым изменяя способность к межбелковому взаимодействию. Например, эпидермальный фактор роста, связывая свой рецептор , активирует его аутокаталитическое фосфорилирование. Фосфорилированный рецептор может связывать адапторный белок GRB2, передающий сигнал на следующий посредник. Далее это может привести к активации сигнального пути Ras-MAPK/ERK . В нём киназа MAPK/ERK (митоген-активируемая протеинкиназа) может далее активировать фактор транскрипции C-myc и изменять функционирование ряда генов, влияющих на клеточный цикл.