Таксономия Флинна
- 1 year ago
- 0
- 0

Эволюцио́нная таксоно́мия (или эволюцио́нная система́тика ) — одно из направлений биологической систематики , наиболее влиятельное в 1950—1970-е гг. и продолжающее сохранять в настоящее время определённое влияние.
Классиками этого направления можно считать Дж. Г. Симпсона и Э. Майра . Такого мнения придерживался, в частности, один из виднейших представителей эволюционной систематики академик А. Л. Тахтаджян , который, ссылаясь на работу , писал, что тот «называет современную эволюционную систематику симпсоновской (Simpsonian), что не лишено оснований, так как именно Simpson (и параллельно Mayr) последовательно развивал дарвиновскую методологию» .
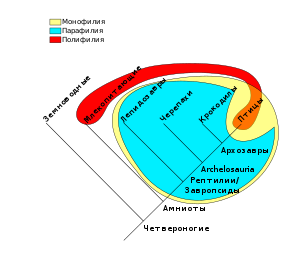
Позитивная программа эволюционной таксономии сформировалась постепенно в ходе дискуссий со сторонниками в 1930—1940-е, фенетики , или нумерической таксономии, в 1950—60-е и кладистами в 1970—80-е гг. Наиболее существенные особенности состояли в том, что — в отличие от сторонников идеалистической морфологии и фенетики — сторонники эволюционной таксономии признавали, что система должна соотноситься с филогенией , но — в отличие от кладистов — они не настаивали на строгом и взаимно-однозначном соответствии между реконструированной схемой филогенетических отношений и классификацией.
В частности, это выражалось в различном отношении к проблеме парафилетических групп. В то время как сторонники эволюционной таксономии были склонны признавать их, считая разновидностью монофилетических, кладисты отвергали парафилетические группы в своих классификациях, настаивая на более узком понимании монофилии .
Говоря о различии между двумя направлениями, А. Л. Тахтаджян отмечал : «Для кладистов филогения сводится к … генеалогии и классификации и оказывается скорее классификацией апоморфных признаков, чем организмов… В отличие от кладистов эволюционная систематика основывается не только на кладистическом анализе, но и на учёте темпов и модусов эволюции отдельных ветвей, степени их дивергенции и уровне специализации».
Для описания различия в подходах эволюционной таксономии и кладистики к понятию «таксона» обсудим кратко различие между апоморфными и плезиоморфными признаками. Апомо́рфный признак — это признак, выделяющий группу видов среди множества других и унаследованный членами группы от их общего предка. Плезиомо́рфный признак — это признак, унаследованный группой видов от их общего предка и характеризующий бо́льшую по объёму группу, содержащую первую группу в качестве своей подгруппы .
Согласно воззрениям кладистов , таксоны могут выделяться только по апоморфным признакам (поскольку группа, при выделении которой берутся в расчёт и плезиоморфные признаки, может оказаться парафилетической ). А. Л. Тахтаджян отмечал в этой связи : «Конечно, значение апоморфных и плезиоморфных признаков для систематики разное. Несомненно, анализ „горизонтальных“ (патристических) взаимоотношений сестринских групп должен быть основан на апоморфных признаках. Однако при выяснении вертикальных взаимоотношений между последовательными ветвлениями кладограммы плезиоморфные признаки приобретают большое, часто решающее значение».
Не менее важно значение плезиоморфных признаков для восстановления облика и строения анцестральных групп. Согласно Л. П. Татаринову , учёт симплезиоморфии имеет большое значение для реконструкции организации предковых организмов, и без этого характеристики предков превращаются в «мозаику ограниченного числа подчас слабо связанных друг с другом апоморфных признаков».
В результате системы, создаваемые по методологиям эволюционной таксономии и кладистики , могут совпадать, а могут и существенно различаться. В качестве примера рассмотрим структуру класса Mammalia ( Млекопитающие ), ограничиваясь лишь (обычно принимаемыми) отрядами , включающими ныне живущие виды. По результатам исследований последних лет , практически все такие отряды (как и сам класс Mammalia ) представляют собой монофилетические ; исключением оказывается отряд Artiodactyla ( Парнокопытные ).
Немонофилетичность Artiodactyla обусловлена тем, что отдельные входящие в него группы образуют монофилетическую группу (получившую название Cetartiodactyla ( Китопарнокопытные )), только при условии, что к ним добавляется группа Cetacea ( Китообразные ), традиционно имеющая ранг отряда. Взаимоотношения между этими группами можно представить в виде следующей кладограммы :
| Cetartiodactyla |
|
||||||||||||||||||||||||
В соответствии с представленной на данной кладограмме филогенией, кладисты не признают Artiodactyla как таксон (поскольку это — парафилетическая группа), и его место в системе занимает таксон Cetartiodactyla (ранг которому не присвоен).
В эволюционной таксономии возможны три подхода. Во-первых, можно сохранить существующую номенклатуру отрядов (парафилетичность Artiodactyla не запрещает считать данную группу полноценным таксоном). Во-вторых, можно (добиваясь монофилетичности всех входящих в Mammalia отрядов) удалить из системы отряд Artiodactyla , придав ранг отряда группам, именуемым ныне Tylopoda , , Ruminantia и Hippopotamidae (с возможной корректировкой некоторых из этих названий). В-третьих, можно придать ранг отряда группе Cetartiodactyla , удалив из системы Artiodactyla и понизив ранг Cetacea до подотряда. Все три подхода не нарушают правил, по которым выделяют таксоны в рамках эволюционной систематики; выбор наиболее предпочтительного из этих подходов предполагает тщательный анализ как дивергенции всех данных групп, так и степени сходства этих групп (и группы Cetacea ).
В настоящее время в мировой практике эволюционная таксономия уступила лидирующее место кладистике . По некоторым оценкам , не менее 90 % работ по систематике на Западе выходит под непосредственным влиянием кладизма. Однако часть учёных (в том числе и весьма крупных — таких, например, как известный специалист в области живых организмов Т. Кавалир-Смит ) продолжает оставаться убеждёнными сторонниками эволюционной таксономии.