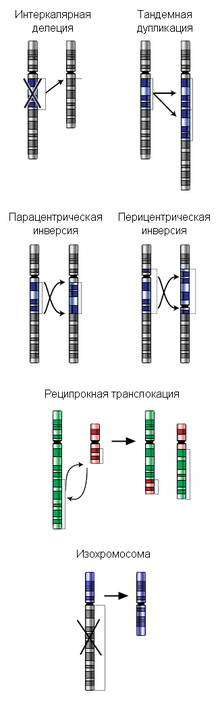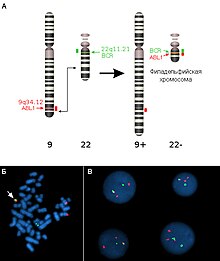
Хромосомные перестройки
- 1 year ago
- 0
- 0
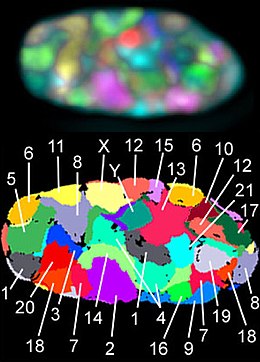
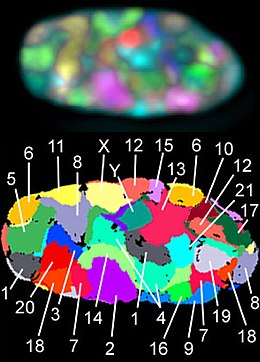
Хромосо́мные террито́рии — отдельные и почти не перекрывающиеся области, которые занимают хромосомы в ядре во время интерфазы клеточного цикла . Хромосомные территории имеют сферическую форму с диаметром от одного до нескольких микрометров . Считается, что территориальная организация хромосом характерна для всех эукариот , хотя есть и исключения, например, дрожжи Saccharomyces cerevisiae . Взаимное расположение хромосомных территорий сильно варьирует от клетки к клетке .
Хромосомы доступны визуальному микроскопическому наблюдению только во время митотического или мейотического деления клеток. Неудивительно поэтому, что после открытия хромосом в XIX веке полагали, что они являются временными образованиями, возникающими в клетке только в момент клеточного деления, а в интерфазе рассыпающимися на отдельные фрагменты. Первым, кто в 1883 году высказал предположение о непрерывном существовании хромосом в течение всего клеточного цикла , был бельгийский эмбриолог Эдуард ван Бенеден . Эта идея была развита в работе австрийского цитолога Карла Рабля « Über Zellteilung » ( нем. «О клеточном делении») (1885). Карл Рабль, наблюдая за процессом деления эпителиальных клеток земноводных Salamandra maculata и Proteus anguinis , обнаружил преемственность расположения хромосом в последовательных делениях. Он высказал гипотезу, что каждая хромосома как отдельная сущность сохраняется в интерфазном ядре, при этом занимает в ядре место, во многом задаваемое позицией в анафазе предшествующего митоза. Кроме этого, Карл Рабль описал характерную конфигурацию хромосом в интерфазном ядре, при которой центромеры хромосом крепятся к ядерной оболочке на одном полюсе ядра, а теломерные участки хромосом — на противоположном полюсе. Подобную организацию хромосом в ядре сейчас называют «конфигурацией Рабля» .
Термин «хромосомная территория» ввёл в научный обиход немецкий эмбриолог Теодор Бовери в работе 1909 года, посвящённой изучению хромосом в бластомерах у лошадиной аскариды Parascaris equorum . По теории непрерывности хромосом, сформулированной Т. Бовери, хромосомы в интерфазе не исчезают, а переходят в деконденсированное состояние, при этом занимая отдельное пространство и не перепутываясь с другими хромосомами .
Экспериментальное подтверждение существования хромосомных территорий было получено значительно позднее — в 80-х годах XX века. Первые косвенные свидетельства того, что хромосомы не являются равномерно распределёнными по ядру, были получены в 1982 году немецким исследователем . Эксперименты Т. Кремера были основаны на повреждении клеточного ядра при помощи тонкого луча УФ -лазера. После такого локального воздействия лишь малая часть хромосом несла следы репарационных изменений. Прямое визуальное доказательство территориальной организации хромосом в ядре было получено Т. Кремером и другими исследователями в 1984-85 годах при помощи гибридизации in situ с радиоактивно-меченой ДНК отдельных хромосом .
Современные представления о расположении в ядре интерфазных хромосом основаны на данных экспериментов по флуоресцентной гибридизации in situ ( англ. fluorescent in situ hybridization, FISH ), в которых в качестве зондов используются тем или иным образом меченные короткие фрагменты индивидуальных метафазных хромосом .
Жёстких правил, которым подчиняется расположение хромосом в интерфазном ядре, нет, однако показано, что гомологичные хромосомы обычно располагаются далеко друг от друга. Кроме того, в ядрах лейкоцитов хромосомы, богатые генами , занимают центр ядра, а бедные генами хромосомы залегают по периферии ядра, ближе к ядерной оболочке . Однако в плоских ядрах фибробластов эта тенденция выражена не так ясно: в ядрах фибробластов все большие хромосомы, как правило, располагаются ближе к ядерной периферии. Химерные хромосомы, которые представляют собой результат транслокации между хромосомами из разных слоёв ядра, занимают промежуточную позицию. Позиция каждой конкретной хромосомы в ядре обозначается путём указания среднего расстояния между центром ядра и центром хромосомной территории, выраженного в процентах радиуса ядра .
Хромосомные территории разделяются особым пространством — интерхроматиновым доменом ( англ. interchromatin domain, ICD ), который служит для перемещения РНК и регуляторных белков . В нём же располагаются ядерные спеклы ( англ. speckles ) и PML-тельца , накапливающие факторы сплайсинга и белок PML соответственно .
Хромосомные территории имеют «губчатую» структуру, благодаря которой внутрь них могут проходить регуляторные белки, а рибонуклеопротеиновые частицы путём простой диффузии перемещаться из мест синтеза в интерхроматиновый домен .
Согласно наиболее обоснованной модели, базовой структурной единицей хромосомных территорий являются домены размером 1 млн п. н. Они представляют собой розетки из 10 и более петель ДНК и соответствуют кластерам репликонов . Хотя первоначально было показано, что активные гены залегают на поверхности хромосомных территорий, дальнейшее изучение показало, что подобной корреляции не существует. Однако в некоторых случаях она прослеживается, например, в случае локуса HoxB мыши .
Функциональное значение выпетливания генов за границы хромосомной территории в данный момент не ясно. Гены, в которых идёт активная транскрипция и которые находятся за пределами хромосомных территорий, часто находятся рядом с PML-тельцами. PML-тельца, помимо PML-белков, могут содержать и другие транскрипционные факторы , поэтому перемещение генов ближе к PML-тельцам может способствовать активации транскрипции. Возможно также, что выход активно экспрессирующихся генов за пределы хромосомных территорий есть простое следствие декомпактизации соответствующего хромосомного домена .
В ходе ответа на повреждения ДНК ( англ. DNA damage response, DDR ) происходит перемещение хромосомных территорий. Так, повреждения в ДНК кожных фибробластов при обработке цисплатином приводят к перемещению хромосомных территорий 12-ой и 15-ой хромосом к центру ядра, а хромосомные территории хромосом 17 и 19 , напротив, перемещаются из центра ядра к его периферии .
| Основное |

|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
|
||||||||||
|
Перестройки и
нарушения |
|||||||||||
|
Хромосомное
определение пола |
|||||||||||
| Методы | |||||||||||