
Полиандрия у насекомых
- 1 year ago
- 0
- 0



Полёт
— один из основных способов передвижения множества
насекомых
различных отрядов, помогающий им искать пищу, полового партнёра для размножения, расселяться и мигрировать, спасаться от хищников.
Насекомые первыми на Земле выработали способность к
полёту
и поднялись в воздух, являясь единственной группой
беспозвоночных
, способной к полёту. Благодаря появлению крыльев насекомые стали более быстрыми и манёвренными, приобрели способность к регулярным
миграциям
; резко усложнилось их поведение. Увеличились возможности для питания и размножения, появились новые способы для избегания врагов. Изучение принципов полёта насекомых представляет большой интерес для сравнительной
физиологии
, систематики, прикладной
энтомологии
и
бионики
.
В течение длительного периода исторического развития система обеспечения полета насекомых прогрессивно развивалась, а их полётные возможности совершенствовались.
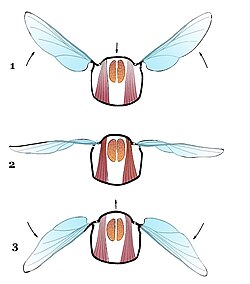
Крылья насекомых (alae) представляют собой пластинчатые выросты покровов, связанные с комплексом обслуживающих их мышц и преобразованных склеритов груди . В типичном случае их две пары: передняя, прикреплённая к среднегруди, и задняя — на заднегруди. Крылья состоят из тонкой крыловой пластинки (membrana), укреплённой на твёрдых жилках (venae), играющих роль прочной основы крыла. Жилки делятся на продольные и поперечные, образующие у более низкоорганизованных групп густую сеть и многочисленные ячейки (cellulae). Поперечные жилки у высших форм сокращаются в числе и иногда исчезают совсем, так как главную поддерживающую роль несут продольные, попеременно выпуклые и вогнутые жилки. Поэтому продольные жилки, хотя и претерпели у высших групп значительную эволюцию, всё же сохранили свою основу .
Крылья насекомых, в отличие от крыльев летающих позвоночных животных, лишены собственной мускулатуры и приводятся в движение сокращением мышц груди.
Продольные спинные мышцы. Мышца соединяет фрагмы и развита у летающих форм. Сокращения данной мышцы приводят к выгибанию тергита вверх, благодаря чему гребная часть крыла опускается. В переднегруди её передний конец крепится к постмаксиллярной складке и участвует также в движении головы. Вторая продольная мышца располагается сбоку от первой и проходит в каждом сегменте от задней фрагмы к скутуму. Основная функция — вспомогательное опускание крыла .
Плейральные мышцы. Целая группа мышц. Наибольшее значение имеют три из них. Первая и вторая, лежащие в пределах эпистерны и сходящиеся кверху к базаларной пластинке. Данная пластинка лежит в мембране, соединяющей крыло с плейритом . При сокращении этих мышц передний край крыла, опускается и изменяется наклон крыла. Также данные мышцы служат для расправления крыла и двигают коксу, и, таким образом, обладают тройной функцией. Третья мышца является антагонистом первых двух и располагается в пределах эпимеры .
Продольные вентральные мышцы. Первая обычно сохраняется у личинок, и отсутствует у имаго . Вторая соединяет фурки стернитов . От переднегрудной фурки идет к тенториуму и осуществляет движение головы. От каждой ветви фурки проходят ещё две мышцы. Первая из них идёт к передней спинальной складке, вторая — к задней. Основная функция данной группы мышц — притягивание сегментов друг к другу .
Дорсо-вентральные мышцы . Данные мышцы являются аналогом кольцевой мускулатуры аннелид. В группу входят шесть мышц, идущих от тергита к стерниту, либо к частям ноги. Первая мышца является терго-стернальной; её функция — притягивание тергита к стерниту, что приводит к поднятию гребной части крыла. Данная мышца наиболее развита у летающих форм. Вторая мышца обладая той же функцией, располагается от передней спинной мышцы коксоподита. Третья и четвёртая мышцы начинаясь на тергите, выполняют двойную функцию: при неподвижной ноге выполняют функцию вспомогательных поднимателей крыла, а также приводят в движение ногу. Пятая мышца располагается от тергита к внутреннему краю вертлуга, и соединена с ним специальным сухожилием. Сокращаясь, данная мышца опускает ногу, и служит вспомогательным поднимателем крыла. Шестая мышца идёт от тергита к мерону, расположенному рядом с коксой, и также обладает двойной функцией .

Полёт насекомых может быть активным и пассивным.
Основной формой передвижения насекомых в воздухе является машущий полёт, который используют все способные к нему крылатые насекомые.
Поступательный машущий полёт насекомых расценивают как манёвренный, экономичный, устойчивый и быстрый, хотя отдельные его показатели могут изменяться у разных групп насекомых.
Крупные насекомые способны к прямолинейному полёту в течение более длительного временного промежутка, чем мелкие. Причём у
жёсткокрылых
данная тенденция выражена гораздо больше, чем у других групп насекомых. Данная тенденция связана со стабилизацией траектории полёта с помощью расставленных в стороны надкрылий.
Траектория полёта большинства видов складывается из различных поворотов, коротких бросков в стороны, петель, неожиданных остановок, резких изменений высоты. Эти изменения сопровождают все манёвры в горизонтальной плоскости и обычны в прямолинейном полёте. Из-за этого траектория полёта часто становится волнистой.
Также полёт насекомых характеризуется выраженной неравномерностью скорости движения. В отдельные моменты насекомое может развивать высокую скорость, а в другие — останавливаться или продолжать полёт с гораздо меньшей скоростью.
В зависающем полёте, в отличие от поступательного, насекомое стремится сохранить своё положение в пространстве неизменным. В англоязычной литературе для обозначения данного типа машущего полёта применяется специальный термин — ховеринг ( англ. hovering ). Он характерен для:

К ховерингу способны бражники, стрекозы , некоторые виды клопов , жуки , а также все двукрылые и перепончатокрылые . Траектория, которую описывают крылья насекомого, находящегося в состоянии зависающего полёта, может быть под разными углами наклонена к горизонтальной плоскости. В зависимости от этого различают три типа зависающего полёта.
При первом типе плоскость взмаха крыла располагается горизонтально, что достигается путём придания телу наклонного или вертикального положения. Крылья насекомого при этом двигаются с большой амплитудой, а их вершины описывают траекторию в форме горизонтальной восьмёрки. В крайней передней и задней точках траектории крыло сильно вращается вокруг своей продольной оси; обе ветви оно проходит с одинаковым по величине углом атаки, причём вектор полной аэродинамической силы направлен вверх. Это так называемый нормальный ховеринг. Его наиболее часто используют в полёте различные насекомые: комары-долгоножки, мухи-сирфиды, пчёлы, бражники, некоторые жуки.
Для ховеринга второго типа характерно горизонтальное расположение тела насекомого, при этом плоскость взмаха крыльев составляет с горизонтом некоторый угол, а амплитуда взмаха крайне мала. Данный способ используют представители двух групп насекомых: сирфиды подсемейства Syrphinae и разнокрылые стрекозы . По наблюдениям, ховеринг второго типа используют также пчелиные и складчатокрылые осы .
Ховеринг третьего типа был описан Ч. Эллингтоном у бабочки капустницы . Крылья двигаются по траектории, перпендикулярной горизонтальной плоскости: вниз — с углом атаки около 90°, вверх — с нулевым. Следует отметить, что такой характер движения крыльев свойствен белянкам и некоторым другим чешуекрылым при поступательном полёте, однако все они не способны к длительному зависанию .
Пассивный полёт по сути является продолжением машущего: крылья не создают тягу, и насекомое движется за счёт энергии, накопленной ранее в активном машущем полёте. При данном типе полёта на насекомое действуют следующие силы:
Движущей силой при планирующем полёте служит составляющая массы тела по направлению перемещения, которая возникает вследствие наклона траектории движения к горизонту. Следовательно, только сравнительно крупные и тяжёлые насекомые способны к пассивному полёту. В среднем площадь крыльев увеличивается при возрастании массы тела, однако не все крупные насекомые способны к пассивному полёту.
Планирующий полёт характерен для насекомых с большой площадью крыльев ( бабочки , стрекозы ). Разогнавшись, они прекращают движения крыльями и некоторое время планируют благодаря инерции . Некоторые насекомые, например, стрекозы, способны не только планировать, но и использовать восходящие потоки воздуха, чтобы подниматься вверх — парящий полёт.
В зависимости от траектории и скорости движения пассивный полёт подразделён на :
Парашютирование обеспечивает наиболее простое взаимодействие крыльев с воздухом и представляет собой медленное снижение по вертикальной траектории. Встречается у насекомых двух групп: подёнок и длинноусых молей рода Adela при полёте в рое. Роящиеся самцы взлетают вертикально, а затем, остановив крылья в приподнятом положении, медленно снижаются.
В основе других типов пассивного полёта лежит принцип
планирующего полёта
. Крыло, обтекаемое потоком воздуха, создаёт в нём возмущения, приводящие к отклонению воздушной массы потока вниз. Подъёмная сила возникает под воздействием воздушного потока на крыло.
Набегающий поток воздуха разделяется крылом на две части — верхнюю и нижнюю, — при этом верхняя часть проходит более длинный путь, чем нижняя, вследствие выпуклости крыла. Исходя из условия о неразрывности потока, скорость потока сверху крыла больше, чем снизу, что вызывает разность давлений — образуется область низкого давления над крылом и область более высокого давления под крылом — что и создаёт подъёмную силу. Последняя прямо пропорциональна площади крыльев и квадрату скорости.
Пикирование представляет собой быстрое снижение с большим углом между направлением полёта и горизонталью. Встречается у насекомых довольно редко: у некоторых чешуекрылых и
саранчовых
. Последние прибегают к нему при изменении высоты полёта в стае, а также в случае опасности. Вспугнутое насекомое прыгает, раскрывая крылья, и поднимается по наклонной траектории машущим полётом, а затем пикирует по такой же наклонной траектории вниз и приземляется.
Например, дальность пикирования
голубокрылой кобылки
(
Oedipoda coerutescens
) с высоты 1 м составляет 1,5—2,0 м
. Такое же поведение (но без прыжка) отмечается у многих видов, обитающих в траве
медведиц
рода
и
совок
. Крылья бабочки при таком пикировании немного отведены назад, что помогает достичь высокой скорости.
Отличие планирования от пикирования заключается в том, что насекомое, набрав скорость в полёте, продолжает движение вперёд без взмахов крыльев, экономя таким образом энергию. Насекомые, использующие планирующий полёт, характеризуются низкими нагрузками на крыло — то есть малым отношением массы тела к площади крыльев.
В свою очередь, парение отличается от планирующего полёта тем, что поток набегает на крыло под большим, чем при планировании,
углом атаки
.
Такие насекомые, как
разнокрылые стрекозы
, некоторые
сетчатокрылые
и
чешуекрылые
используют для парящего полёта также восходящие потоки воздуха. Так, среди чешуекрылых способностью к парению обладают представители семейств:
данаиды
,
Nymphalidae
,
Papilionidae
,
Pieridae
,
Morphinae
,
Uraniidae
.
Мелкие виды из семейства
нимфалиды
для парения используют термические потоки, образующиеся у самой земли, чернушка
может летать кругами над дорогой, от которой поднимаются нагретые потоки воздуха. Более крупные виды парят выше, часто на уровне крон деревьев.
Ещё более приспособленными к парению являются представители семейства
парусников
.
Обыкновенный аполлон
(
Parnassius apollo
) может длительное время парить над склонами горных ущельев, корректируя траекторию полёта взмахами крыльев и меняя высоту.
Атрофанейра алкиной
при парящем полёте летает на уровне крон, используя прерывистые взмахи — при этом передние крылья распластаны, а задние приподняты и сложены «лодочкой».
Повышение устойчивости при парящем полёте достигается длинными хвостовидными выростами задних крыльев.
Особенно большой длины эти выросты достигают у сетчатокрылых, например,
Nemoptera sinuata
, и некоторых видов бабочек из семейства
Сатурнии
(
Actias artemis
,
Coscinocera hercules
,
Argema mimosae
). Их задние крылья имеют длинные хвостовидные выросты, закрученные в направлении, противоположном концевым завихрениям, и совпадающем с направлением вращения вихревых потоков у внутреннего края. Хвостовидные выросты при планирующем полёте сближены и направлены строго назад.
Все виды, способные к парению, также используют и планирование . Характерным признаком приспособления к последнему служит вытянутость вершин передних крыльев. К планированию часто прибегают нимфалиды , парусники , данаиды и другие чешуекрылые .
Полёт насекомых контролируется рефлекторно и поддерживается стимуляцией механочувствительных сенсилл у оснований крыльев. Отклонения от ориентации во время полёта могут быть описаны как вращения вокруг трёх основных пространственных осей:
Также ориентация поддерживается за счёт зрительной информации, частично — информации от чувствительных органов у сочленения головы и груди. У двукрылых — частично от жужжалец .
У стрекоз, например, имеется три подушечки волосковых сенсилл с каждой стороны сочленения головы и груди, которые способны детектировать угловые ускорения. В полёте голова поддерживается в своём положении зрительной информации от глаз. При изменениях и нарушениях положения тела, сенсиллы волосковых подушечек воспринимают данные изменения. Две пары крыльев, при этом работая рефлекторно, восстанавливают соосность расположения головы и тела. Такой же механизм существует и у большинства других летающих насекомых.
У двукрылых задняя пара крыльев модифицируется в гантелевидные жужжальца с многочисленными колоколовидными сенсиллами у их оснований. Жужжальца колеблются синхронно с движениями крыльев на 180 градусов, с частотой от 100—500 Гц. Колоколовидные сенсиллы воспринимают различные силы, действующие на сочленении жужжалец с грудью при кабрировании, рысканьи и закрутке. Экспериментальное удаление жужжалец приводило к нарушениям рефлекторного компенсаторного контроля мышц крыльев.
Скорость перемещения насекомых в первую очередь зависит от ветра. Различают скорость абсолютную (воздушную) — скорость полёта насекомого во время штиля, и скорость относительную (земную), которая зависит от скорости и направления ветра.
Путём сложения векторов скорости ветра и воздушной скорости объекта можно определить земную скорость последнего. При попутном ветре насекомое достигает более высокой скорости перемещения относительно элементов ландшафта. Так, бабочка-монарх при попутном ветре может лететь со скоростью более 14 м/сек.
В среднем скорость полёта насекомых колеблется у различных видов от нескольких сантиметров в секунду до 15 м/сек. Максимальная скорость полёта характерна для таких насекомых, как
разнокрылые стрекозы
,
бражники
и
.
Манёвренность можно определить, как скорость изменения направления движения. Ускорение, развиваемое в полёте, достигает максимальных значений у двукрылых: журчалок и жужжал. За ними следуют стрекозы . В зависимости от характера полёта стрекоза рода Libellula в состоянии менять скорость так быстро, что ускорение составляет 2,5g. Движение с наибольшим ускорением, равным 9g, отмечено у стрекоз семейства Corduliidae . Наименьшая скорость маневрирования среди стрекоз свойственна представителям семейства Calopterygidae : поворот на 90° занимает у них 0,5 сек., но зато эти повороты могут следовать один за другим без перерыва, образуя каскад .
Активный полёт осуществляется за счёт энергии сокращений мышц, пассивный — за счёт использования энергии внешней среды и массы тела насекомого.
|
|
Этот раздел
не завершён
.
|
|
|
Этот раздел
не завершён
.
|
|
|
Этот раздел
не завершён
.
|
В зависимости от скорости, выполнения определённых манёвров, использования ховеринга и других особенностей поведения в воздухе можно выделить 10 основных типов полёта, встречающихся у современных насекомых . Между типами полёта нет чётких границ, и существуют многочисленные переходы от одного к другому.
1-й тип (рыскающий) свойствен относительно примитивным крылатым насекомым: подёнкам , веснянкам , вислокрылкам , скорпионницам , некоторым ручейникам . Полёт часто непродолжителен: взлетевшее насекомое стремится как можно скорее приземлиться. Для него характерна изломанная траектория, что связано с частой сменой высоты; при этом изменение направления движения на коротком отрезке пути не сопровождается поворотом продольной оси тела в сторону перемещения. Перелёт первичных зубатых молей с цветка на цветок происходит не по кратчайшему пути, а из-за рыскающих движений по сложной траектории. Характерен полёт по спирали (мелкие подёнки, ручейники, первичные моли и другие). Скорость полёта, как правило, низкая. Частая смена высоты вызвана неравномерностью работы крыльев.

2-й тип свойствен летающим среди густой растительности хищникам ( верблюдки , некоторые сетчатокрылые ). При таком типе полёта также отсутствует поворот продольной оси тела в направлении движения при кратковременном его изменении. Чрезвычайно характерен полёт вверх, при котором тело ориентировано вертикально. Эта особенность, а также сложные манёвры, выполняемые на невысокой скорости, позволяют отличить этот тип полёта от предыдущего.
3-й тип характеризуется сравнительно низкой скоростью перемещения в любом направлении без поворота тела. Он свойствен равнокрылым стрекозам , которые обладают способностью мгновенно менять направление движения, им же свойствен ховеринг. Подъём эти насекомые, в отличие от златоглазки , осуществляют мгновенным рывком, сохраняя горизонтальное положение тела.
4-й тип свойствен разнокрылым стрекозам , для которых характерен ховеринг второго типа. Отличительная черта полёта разнокрылых стрекоз — способность к быстрым поворотам тела практически в одной точке пространства: летящая стрекоза резко останавливается и в ховеринге поворачивается в нужном направлении, после чего продолжает полёт. Угол поворота может быть 90° и больше. Набор высоты происходит очень быстро и без разворота тела.
5-й тип близок к третьему, но отличается значительно более высокой скоростью перемещений. Он свойствен летающим среди травянистой растительности двукрылым: толкунчики , ктыри , бекасницы , журчалки и другие. Для этих насекомых характерны частые зависания и резкие броски в стороны, вперёд, назад, вверх. При охоте или отыскивании партнёра для спаривания в зарослях необходима высокая манёвренность, но без значительного смещения в пространстве. Этим целям хорошо отвечает полёт с использованием ховеринга. У некоторых журчалок развилась более прогрессивная форма ховеринга — второго типа.

6-й тип близок к пятому и свойствен большинству перепончатокрылых и многим двукрылым ( слепни , ). Эти насекомые также часто используют ховеринг, но всё же реже, чем вышерассмотренные двукрылые. В то же время такой тип полёта чаще всего встречается при быстрых и дальних перемещениях. Изменение направления движения сопровождается быстрым поворотом тела без остановки.
7-й тип свойствен бражникам. Наиболее близок он к предыдущему, но используется при более дальних перелётах в поисках нужного растения. Из-за крупных размеров и большого веса бражники вынуждены питаться, зависая перед цветком. Приспособление к ховерингу и скоростному полёту произошло в ущерб манёвренности — траектория их полёта более прямолинейна, чем у перепончатокрылых.
8-й тип характеризуется наибольшей скоростью и манёвренностью и быстрой сменой виражей. Сходный характер полёта наблюдается у некоторых короткоусых двукрылых.
9-й тип свойствен бабочкам : части булавоусых, пяденицам , павлиноглазкам и другим. За каждый взмах крыльев насекомое пролетает сравнительно большое расстояние, а периодические изменения высоты и направления придают полёту характер порхающего. Скорость полёта высокая.
10-й тип объединяет полёт таких насекомых, как прямокрылые , жуки и другие. Изменение направления движения сопровождается поворотом продольной оси тела, но манёвренность в целом низкая. Скорость полёта может быть как низкая, так и высокая. Очевидно, что дальнейшее изучение характера полёта различных групп насекомых позволит дифференцировать данный тип. Однако можно говорить лишь о некотором своеобразии полёта жуков, отличающегося низкими манёвренностью и скоростью, хотя последний показатель возрастает у крупных видов.



