
Философия сознания
- 1 year ago
- 0
- 0

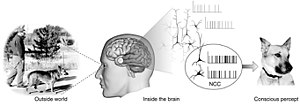
Нейронные корреляты сознания ( англ. neural correlates of consciousness , NCC) — механизмы взаимодействия между психическими и нейронными состояниями; составляют минимальный набор нейронных событий и механизмов, достаточный для конкретного сознательного восприятия . Для обнаружения нейронных коррелятов нейробиологи используют эмпирические подходы , то есть нейронные изменения, которые регулярно коррелируют с конкретным опытом .
Наука о сознании должна объяснить точную взаимосвязь между субъективными психическими состояниями и состояниями мозга, природу взаимосвязи между сознательным разумом и электрохимическими взаимодействиями в мозге ( проблема разум-тело ). Прогресс в нейропсихологии произошел благодаря сосредоточению внимания на теле (мозге), а не на разуме. В этом контексте можно рассматривать нейронные корреляты сознания, а сознание можно рассматривать как свойство неопределенной сложной , адаптивной и тесно взаимосвязанной биологической системы .
Обнаружение и характеристика нейронных коррелятов не предлагает каузальной теории сознания, которая могла бы объяснить, как определенные системы что-то переживают, то есть не решают так называемую трудную проблему сознания , но понимание NCC может быть шагом к каузальной теории. Большинство нейробиологов предполагают, что факторы, порождающие сознание, должны быть найдены на нейронном уровне, управляемом классической физикой. Предлагаются также теории квантового сознания , основанные на квантовой механике .
В ходе изучения нейронных коррелятов сознания (NCC) возникли следующие вопросы: каковы основные свойства NCC; что общего между NCC для зрения и для слуха? Будет ли NCC задействовать все
пирамидальные нейроны
коры в любой данный момент времени или только подмножество проекционных клеток дальнего действия в лобных долях, которые проецируются на сенсорную кору в задней части? И др.
.
Есть два общепринятых, но различных определения термина'«сознание»: одно включает возбуждение и состояния сознания, а другое — содержание сознания и состояния сознания . Чтобы осознать что-либо, мозг должен находиться в относительно высоком состоянии возбуждения (иногда называемого бдительностью ), будь то в состоянии бодрствования или в фазе быстрого сна , которое ярко переживается во сне, хотя обычно не запоминается. Уровень возбуждения мозга колеблется в соответствии с циркадным ритмом , но на него могут влиять недостаток сна, употребление наркотиков и алкоголя, физические нагрузки и т. д. Возбуждение может быть измерено поведенчески по амплитуде сигнала, который запускает некоторую заметную реакцию (например, уровень звука, необходимый для того, чтобы вызвать движение глаз или поворот головы в сторону источника звука). Для оценки уровня возбуждения пациентов используются системы оценки, такие как шкала комы Глазго .
Клиницисты говорят о нарушениях сознания как о « коматозном состоянии », « стойком вегетативном состоянии » (PVS) и « состоянии минимального сознания » (MCS). Здесь «состояние» относится к разным уровням внешнего/физического сознания, от полного отсутствия в коме, стойкого вегетативного состояния и общей анестезии до флуктуирующей и ограниченной формы сознательного ощущения в состоянии минимального сознания, такого как ходьба во сне или во время сложного парциального эпилептического припадка . Набор сознательных состояний или переживаний, доступных пациенту в состоянии минимального сознания, сравнительно ограничен. При смерти мозга возбуждения нет, однако неизвестно, прервана ли субъективность опыта, а не его наблюдаемая связь с организмом. Исследования показали, что у вегетативных пациентов, которые предположительно находятся без сознания, части коры все еще активны . Однако, эти области, по-видимому, функционально не связаны с ассоциативными областями коры, активность которых необходима для осознания.
Потенциальное богатство сознательного опыта увеличивается от глубокого сна к сонливости и полному бодрствованию, что можно количественно определить, используя понятия из теории сложности . По мере увеличения поведенческого возбуждения увеличивается диапазон и сложность возможного поведения. Тем не менее, в фазе быстрого сна наблюдается характерная атония , низкое двигательное возбуждение и человек с трудом просыпается, но при этом сохраняется высокая метаболическая и электрическая активность мозга и яркое восприятие.
Многие ядра с отчетливыми химическими сигнатурами в таламусе , среднем мозге и Варолиевом мосту должны функционировать, чтобы субъект находился в достаточном состоянии возбуждения мозга и мог вообще что-либо испытывать. Таким образом, эти ядра относятся к факторам, обеспечивающим сознание. И наоборот, вполне вероятно, что конкретное содержание любого конкретного сознательного ощущения опосредовано конкретными нейронами коры и связанными с ними структурами, включая миндалевидное тело , таламус , клауструм и базальные ядра .
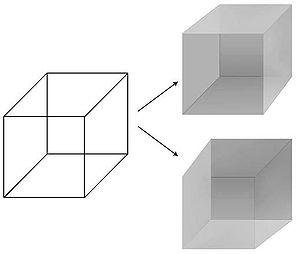
Возможность точного манипулирования визуальными восприятиями во времени и пространстве сделала зрение предпочтительным объектом при изучении NCC. Психологи усовершенствовали для этого ряд техник — маскирование, бинокулярное соперничество , непрерывное подавление вспышек, слепоту, вызванную движением, слепоту к переменам, слепоту по невниманию . В иллюзии восприятия физический стимул остается фиксированным, в то время как восприятие колеблется. Самый известный пример — куб Неккера , 12 линий которого можно воспринимать одним из двух способов.
Иллюзия восприятия, которую можно точно контролировать, это бинокулярное соперничество . Маленькое изображение, например горизонтальная решетка, предъявляется левому глазу, а другое изображение, например вертикальная решетка, показывается в соответствующем месте правого глаза. Несмотря на постоянный визуальный стимул, испытуемые видят, как горизонтальная решетка каждые несколько секунд сменяется вертикальной. Мозг не позволяет одновременно воспринимать оба изображения .
ФМРТ продемонстрировала нормальные проявления мозговой активности у пациента в вегетативном состоянии после тяжелой черепно-мозговой травмы, когда его попросили представить игру в теннис или посещение комнат в его доме . Дифференциальная томография головного мозга пациентов с такими глобальными нарушениями сознания (включая акинетический мутизм ) показывает, что дисфункция в обширной сети коры, включая медиальные и латеральные префронтальные и теменные ассоциативные области, связана с глобальной потерей сознания . Нарушение сознания при эпилептических припадках также сопровождалось снижением мозгового кровотока в лобной и теменной ассоциативной коре и увеличением срединных структур, таких как медиодорсальный таламус .
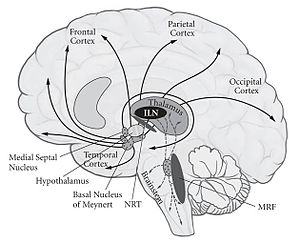
Относительно локальные двусторонние повреждения срединных (парамедианных) подкорковых структур также могут вызывать полную потерю сознания . Таким образом, эти структуры обеспечивают и контролируют возбуждение мозга (определяемое метаболической или электрической активностью) и являются необходимыми нервными коррелятами. Одним из таких примеров является гетерогенное скопление более двух десятков ядер на каждой стороне верхней части ствола мозга (варолиев мост, средний мозг и задний гипоталамус), которые в совокупности называются ретикулярной активирующей системой (РАС). Их аксоны расходятся по всему мозгу.
Эти ядра, — трехмерные скопления нейронов со своей собственной цитоархитектурой и нейрохимической идентичностью, — высвобождают различные , такие как ацетилхолин , норадреналин / норэпинефрин , серотонин , гистамин и орексин / гипокретин , что позволяет контролировать возбудимость таламуса и переднего мозга, опосредуя чередование периодов бодрствования и сна, а также общий уровень поведенческого и мозгового возбуждения.
Возбудимость таламуса и переднего мозга может восстановиться после травмы, и сознание может вернуться . Еще одним фактором, обеспечивающим сознание, являются пять или более ядер таламуса (ILN). Они получают данные от многих ядер ствола мозга и передают сигналы в базальные ганглии и, — более распределенным образом, — в слой I большей части неокортекса. Сравнительно небольшие (1 см 3 или меньше) двусторонние поражения ILN таламуса полностью отключают сознание .
Многие действия в ответ на сенсорные сигналы являются быстрыми, преходящими, стереотипными и бессознательными . Их можно рассматривать как корковые рефлексы, и они характеризуются быстрыми и несколько стереотипными реакциями, которые могут принимать форму довольно сложного автоматизированного поведения, как это наблюдается, например, при сложных парциальных эпилептических припадках. Этим автоматическим реакциям, иногда называемым поведением зомби , можно противопоставить более медленный, универсальный сознательный режим, который работает с более широкими, менее стереотипными аспектами сенсорных входов (или их отражением) и требуется время, чтобы определиться с подходящими мыслями и ответами. Без такого режима сознания потребовалось бы огромное количество различных режимов зомби, чтобы реагировать на необычные события.
Особенность, которая отличает людей от большинства животных, заключается в том, что мы не рождаемся с обширным репертуаром поведенческих программ, которые позволили бы нам выжить самостоятельно (см. « физиологическая недоношенность »). Чтобы компенсировать это, у нас есть непревзойденная способность учиться, т. е. сознательно усваивать такие программы путем имитации или исследования. После сознательного усвоения и достаточной тренировки эти программы могут стать автоматизированными до такой степени, что их выполнение происходит за пределами нашего сознания. Возьмем, к примеру, невероятную мелкую моторику, проявляемую при исполнении фортепианной сонаты Бетховена, или сенсомоторную координацию, необходимую для езды на мотоцикле по извилистой горной дороге. Такое сложное поведение возможно только потому, что достаточное количество задействованных подпрограмм может быть выполнено с минимальным или даже приостановленным сознательным контролем. На самом деле сознательная система может до некоторой степени вмешиваться в работу этих автоматизированных программ .
С эволюционной точки зрения полезно иметь как автоматизированные поведенческие программы, которые могут быстро выполняться стереотипным и автоматизированным образом, так и несколько более медленную систему, дающую время для обдумывания и планирования более сложного поведения. Этот последний аспект может быть одной из основных функций сознания. Некоторые философы считают, что сознание не является необходимым для какого-либо функционального преимущества в эволюционных процессах . Они утверждают, что никто не дал причинно-следственного объяснения того, почему функционально эквивалентный бессознательный организм (то есть философский зомби ) не может достичь тех же преимуществ выживания, что и сознательный организм. Если эволюционные процессы слепы к разнице между функцией F , выполняемой сознательным организмом О и бессознательным организмом О* , неясно, какое адаптивное преимущество может обеспечить сознание . В результате экзистенциальное объяснение сознания получило поддержку у некоторых теоретиков, которые постулируют, что сознание не развилось как адаптация, а было экзаптацией , возникшей в результате других событий, таких как увеличение размера мозга или перестройка коры . Сознание в этом смысле сравнивают со слепым пятном в сетчатке, где оно не является адаптацией сетчатки, а просто побочным продуктом того, как аксоны сетчатки были соединены проводами . Несколько ученых, в том числе Пинкер , Хомский , Эдельман и Лурия , указали на важность появления человеческого языка как важного регулирующего механизма обучения и памяти в контексте развития сознания высшего порядка.
Представляется возможным, что зрительные зомби-режимы в коре в основном используют дорсальный поток в теменной области . Однако теменная активность может влиять на сознание, вызывая воздействие внимания на вентральный поток, по крайней мере, при некоторых обстоятельствах. Сознательный режим зрения во многом зависит от ранних зрительных областей (за пределами V1) и особенно от вентрального потока.
Казалось бы, сложная визуальная обработка (например, обнаружение животных в естественных, загроможденных сценах) может быть выполнена корой головного мозга человека в течение 130–150 мс , то есть времени слишком короткого для движений глаз и сознательного восприятия. Кроме того, рефлексы, такие как окуловестибулярный рефлекс, происходят в еще более коротких временных масштабах. Вполне вероятно, что такое поведение объясняется волной спайковой активности, которая проходит от сетчатки через V1 в V4, IT и префронтальную кору, пока не воздействует на моторные нейроны в спинном мозге, которые контролируют нажатие пальцем как в типичном лабораторном эксперименте. Гипотеза о том, что основная обработка информации является прямой, напрямую подтверждается коротким временем (около 100 мс), необходимым для появления избирательного ответа в IT-клетках.
И наоборот, считается, что сознательное восприятие требует более устойчивой реверберационной нейронной активности, скорее всего, через глобальную обратную связь от лобных областей неокортекса обратно к сенсорным областям коры , которая со временем накапливается, пока не превысит критический порог. В этот момент устойчивая нервная активность быстро распространяется на теменную, префронтальную и переднюю поясную кору, таламус, клауструм и связанные с ними структуры, которые поддерживают кратковременную память, мультимодальную интеграцию, планирование, речь и другие процессы, тесно связанные с сознанием. Конкуренция препятствует одновременному и активному представлению более одного или очень небольшого числа восприятий. Это основная гипотеза теории глобального рабочего пространства сознания .
Короче говоря, в то время как быстрая, но преходящая нейронная активность в таламо-кортикальной системе может поддерживать сложное поведение без сознательного ощущения, предполагается, что сознание требует устойчивой, но хорошо организованной нейронной активности, зависящей от дальнодействующей кортико-кортикальной обратной связи.
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
{{
cite journal
}}
: Википедия:Обслуживание CS1 (множественные имена: authors list) (
ссылка
)




























