Наполеон I
- 1 year ago
- 0
- 0
Рецептивное поле
(
англ.
receptive field
) сенсорного
нейрона
— участок с
рецепторами
, которые при воздействии на них определённого
стимула
приводят к изменению
возбуждения
этого нейрона.
Концепция рецептивных полей может быть применима ко всей
нервной системе
. Если множество сенсорных рецепторов образует
синапсы
c единственным нейроном, они совместно формируют рецептивное поле этого нейрона. Например, рецептивное поле
ганглионарной
(ганглиозной) клетки
сетчатки
глаза
представлено
(
палочками
или
колбочками
), а группа ганглионарных клеток, в свою очередь, создаёт рецептивное поле для одного из нейронов
мозга
. В итоге к одному нейрону более высокого
синаптического
уровня сходятся импульсы от многих
; и этот процесс называется
.
В качестве рецептивных полей слуховой системы могут рассматриваться части слухового пространства ( англ. auditory space ) или диапазоны слуховых частот . Лишь немногие исследователи трактуют слуховые рецептивные поля как определённые участки сенсорного эпителия , например, группы волосковых клеток спирального органа улитки внутреннего уха млекопитающих .
Рецептивные поля соматосенсорной системы — это участки кожи или внутренних органов . Размеры рецептивных полей неодинаковы и зависят от типа механорецепторов .
Большое рецептивное поле нейрона позволяет отслеживать изменения на большей площади чувствительной поверхности, но обеспечивает меньшую разрешающую способность ощущения . Таким образом, пальцы , которые должны осязать тонкие детали, имеют множество плотно расположенных (до 500 на 1 см 3 ) механорецепторов с маленькими рецептивными полями (около 10 мм 2 ), тогда как спина , бёдра и голени имеют меньшее количество рецепторов, объединённых в большие рецептивные поля. Как правило, в центральной части большого рецептивного поля имеется одно «горячее пятно», стимуляция которого вызывает наиболее интенсивный ответ.
Нейроны коры головного мозга , связанные с тактильными ощущениями, имеют рецептивные поля на коже , размеры и расположение которых могут изменяться по мере накопления человеком индивидуального опыта или вследствие повреждения сенсорных (афферентных) нервных волокон . В основном эти нейроны имеют относительно большие рецептивные поля — гораздо большего размера, чем у нейронов спинномозгового узла (спинального ганглия) . Тем не менее, благодаря особым механизмам возбуждения и торможения в рецептивных полях, улучшающим пространственное разрешение, эти кортикальные нейроны способны распознавать тонкие детали.
Рецептивные поля
зрительной системы
можно считать частями зрительного
пространства
(
англ.
visual space
). Например, в качестве рецептивного поля одной
фоторецепторной клетки
можно рассматривать
конус
, охватывающий все возможные направления, с которых эта клетка способна воспринимать
свет
. Его вершина находится в центре
хрусталика
, а основание — в
бесконечности
зрительного пространства. Но традиционно
зрительные
рецептивные поля изображаются на
плоскости
— как
круги
,
квадраты
,
прямоугольники
… Подобные изображения по сути являются сечениями конуса, отвечающего рецептивному полю одной специфической
клетки
, плоскостью, в которой исследователь предъявлял конкретный визуальный стимул. Рецептивные поля
бинокулярных нейронов
первичной
зрительной коры
(или
стриарной области
—
поле Бродмана
17, зрительная зона V1) не уходят в оптическую бесконечность, а ограничены определённым расстоянием от
точки
, в которую направлен взгляд — «
точки фиксации глаз
» (
См.
—
англ.
Panum's area
).
Рецептивные поля нейронов зачастую определяются как области сетчатки, освещение которых изменяет возбуждение конкретного нейрона. Для ганглионарных (ганглиозных) клеток сетчатки эта область включает все фоторецепторы — палочки или колбочки одного глаза, связанные с конкретной ганглионарной клеткой посредством синаптических контактов с
биполярными
,
и
(амакриновыми) клетками. Для бинокулярных нейронов
зрительной коры
рецептивные поля определяются как совокупность соответствующих областей сетчаток правого и левого глаза. Эти области могут быть закартированы по отдельности в каждой сетчатке (при закрывании другого глаза), но полностью связь каждой из областей с возбуждением исследуемого нейрона обнаруживается только в том случае, когда открыты оба глаза.
Хьюбел
и
Визель
(например, Hubel, 1963) развили теорию о том, что рецептивные поля клеток каждого уровня зрительной системы формируются синаптическими соединениями с клетками более низкого иерархического уровня этой системы. В этом случае небольшие и просто устроенные рецептивные поля могут комбинироваться, формируя обширные и сложные рецептивные поля. Позднее
нейробиологи
усовершенствовали эту относительно простую концепцию, допустив, что нейроны низших уровней зрительной системы связаны обратными эфферентными связями с нейронами более высоких уровней.
В настоящее время составлены карты рецептивных полей для клеток всех уровней зрительной системы — от фоторецепторов и ганглионарных (ганглиозных) клеток сетчатки до нейронов латерального (наружного) коленчатого тела , первичной и экстрастриарной зрительной коры. Исследования, основанные лишь на ощущениях, не могут дать полной картины для понимания феномена зрения, поэтому здесь, так же, как и при изучении мозга, должны применяться электрофизиологические методы — тем более, что в эмбриогенезе млекопитающих сетчатка возникает в процессе дальнейшей дифференциации латеральных выпячиваний промежуточного мозга (так называемых глазных пузырей ).

Каждая ганглионарная (ганглиозная) клетка или оптическое нервное волокно (
англ.
optic nerve fiber
) порождает рецептивное поле, расширяющееся по мере возрастания интенсивности освещения. Если размер поля максимален, то свет на его периферии интенсивнее, нежели в центре, отражая то, что некоторые синаптические пути предпочтительнее других.
Организация рецептивных полей ганглиозных клеток, составленных из входов многих палочек или колбочек, позволяет обнаруживать
контраст
, что используется для выявления краевых частей наблюдаемых объектов. Каждое рецептивное поле подразделяется на две части: центральный диск — «
центр
» и концентрическое кольцо — «
периферию
»; каждая из этих частей реагирует на свет противоположным образом. Так, если освещение центра рецептивного поля увеличивает возбуждение конкретной ганглиозной клетки с так называемым
on-центром
(см. далее), то воздействие света на периферию этого же поля оказывает тормозящее воздействие на эту ганглиозную клетку.
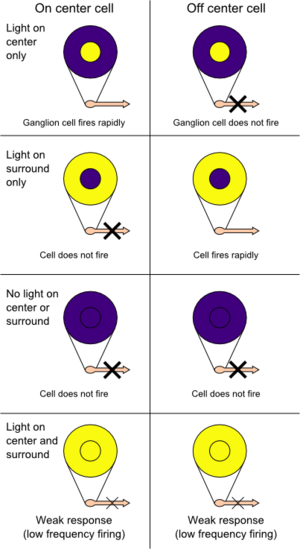
Существует два основных типа ганглиозных клеток: с «
on-центром
» и «
off-центром
». Клетка с
on-центром
возбуждается при освещении центра и тормозится при освещении периферии её рецептивного поля. Реакция на свет клетки с
off
-центром диаметрально противоположная. Кроме того, у млекопитающих имеются клетки промежуточного (
on-off
) типа, которым свойственна кратковременная реакция на освещение по
on
-типу и на затенение по
off
-типу.
Освещение центральной части рецептивного поля приводит к
деполяризации
и возрастанию возбуждения нейрона (например, ганглионарной клетки) с
on
-центром, освещение периферии рецептивного поля приводит к
гиперполяризации
и торможению этого нейрона, а одновременная световая стимуляция и центра, и периферии рецептивного поля вызывает слабую активацию (вследствие суммации эффектов, связанных с реакциями центральной и периферической частей рецептивного поля). Ганглионарная клетка (или другой нейрон) с
off
-центром возбуждается при световой стимуляции периферии и тормозится при освещении центра своего рецептивного поля (см. рисунок).
Фоторецепторы, которые включены в состав рецептивных полей нескольких ганглиозных клеток, способны как возбуждать, так и тормозить постсинаптические нейроны (
англ.
postsynaptic neurons
), поскольку они высвобождают
нейротрансмиттер
глутамат
на своих синапсах, что может способствовать как деполяризации, так и гиперполяризации мембранного потенциала клетки, в зависимости от того, какие именно
ионные каналы
открываются нейротрансмиттером. Организация рецептивного поля по принципу
центр-периферия
позволяет ганглиозным клеткам передавать
информацию
не только о том, освещены ли фоторецепторные клетки, но также и о различиях в параметрах возбуждения подобных клеток, расположенных в центре и на периферии рецептивного поля. Последнее даёт возможность ганглиозным клеткам посылать нейронам более высоких синаптических уровней информацию о контрастности изображения.
Размер рецептивного поля влияет на пространственную частоту (
англ.
spatial frequency
) визуальной информации: небольшие рецептивные поля активируются сигналами с высокими пространственными частотами и тонкой детализацией изображения; большие рецептивные поля — сигналами с низкими пространственными частотами и плохой детализацией.
Рецептивные поля ганглиозных клеток сетчатки передают информацию о дискретности распределения света, падающего на сетчатку, а это зачастую позволяет обнаруживать краевые части визуальных объектов. При адаптации к темноте инактивируется периферийная зона рецептивного поля, но его активная часть, а следовательно, площадь суммации сигналов и совокупная чувствительность, могут реально возрасти вследствие ослабления взаимного горизонтального торможения центра и периферии рецептивного поля.
Как правило, рецептивные поля лучше реагируют на движущиеся объекты — такие как светлое или тёмное пятно, пересекающее поле от центра к периферии (или в противоположном направлении), а также на контуры объектов — вследствие нарушения равномерности в распределении света по поверхности поля. Диаметр центральной части рецептивного поля ганглионарной клетки сетчатки совпадает с протяжённостью её
дендритов
, тогда как площадь периферии рецептивного поля определяется амакринными клетками, устанавливающими связь данной ганглионарной клетки со множеством биполярных клеток. Кроме того, амакринные клетки могут не допускать передачи сигналов в ганглионарную клетку от периферии её рецептивного поля, тем самым усиливая доминирование реакции центра рецептивного поля («включённый центр и выключенная периферия» —
англ.
“on-center, off-periphery”
). Ганглионарная клетка сетчатки
кролика
возбуждается при движении светового пятна в «предпочитаемом» (
англ.
"preferred"
) направлении и не реагирует, если направление является противоположным («нулевым»,
англ.
"null"
).
Ганглиозные клетки, способные различать направление движения, найдены также в сетчатке кошки, земляной белки, голубя. Считается, что обнаруженные свойства рецептивных полей ганглиозных клеток связаны с особенностями сложных механизмов торможения, действующих в сетчатке.
На более высоких уровнях зрительной системы группы ганглионарных (ганглиозных) клеток формируют рецептивные поля нейронов подкоркового зрительного центра —
латерального (наружного) коленчатого тела
. Рецептивные поля напоминают таковые ганглионарных клеток, с антагонистической системой «центр-периферия»; здесь также имеются нейроны с
on-
или
off-
центрами (приблизительно в равном количестве).
Рецептивные поля нейронов зрительной зоны коры крупнее по размерам и имеют большую избирательность по отношению к визуальным стимулам, нежели ганглиозные клетки сетчатки или нейроны латерального коленчатого тела. Хьюбел и Визель (например, Hubel, 1963) подразделили рецептивные поля корковых нейронов зрительной системы на «простые» , «сложные» и «сверхсложные» . «Простые» рецептивные поля имеют удлинённую форму, к примеру, с центральной эллипсовидной зоной возбуждения и антагонистической зоной торможения по периферии эллипса. Либо они могут быть почти прямоугольными; при этом одна из длинных сторон прямоугольника является зоной возбуждения, а другая — антагонистической зоной торможения. Изображения, активирующие нейроны этих рецептивных полей, должны быть ориентированы определённым образом. Чтобы возбудить нейрон со «сложным» рецептивных полем, световому стимулу в виде полоски недостаточно быть правильно ориентированным — нужно ещё и двигаться, причём в строго определённом направлении. Для активации корковых нейронов со «сверхсложными» рецептивными полями зрительному стимулу в виде полоски необходимо обладать всеми вышеперечисленными свойствами, и к тому же длина этой полоски должна быть строго определённой.
Экстрастриарная зрительная кора ( поля Бродмана 18 и 19) находится за пределами первичной зрительной коры . Здесь нейроны могут иметь очень большие рецептивные поля, и для их активации могут потребоваться очень непростые изображения. Например, рецептивные поля нейронов нижневисочной извилины (англ. ), пересекают среднюю линию зрительного пространства, и эти нейроны активируются такими сложными визуальными образами, как радиальная решётка или кисти рук. Также было обнаружено, что нервные клетки вентральной поверхности веретеновидной извилины (на границе между затылочной и височной долями), где находится так называемая , реагируют, в основном, на изображения лиц . Это важное открытие было получено с помощью технологии функциональной магнитно-резонансной томографии . Позднее оно было подтверждено на уровне исследования нервных клеток . Подобным способом проводятся поиски других специфических зон зрительной коры; например, имеются относительно недавние публикации, полагающие, что так называемая парагиппокампальная навигационная зона (англ. ) может быть отчасти специализирована к распознаванию зданий. Кстати, в одном из последних исследований высказывается предположение, что «зона распознавания лиц» веретеновидной извилины, возможно, не только выполняет функцию, отражённую в её наименовании, но и вообще служит для различения отдельных частей целого.
|
|
|
|---|