Interested Article - Клетка Реншоу

- 2020-12-11
- 1
Клетки Реншоу (КР) ( англ. Renshaw cells ) — тормозные вставочные нейроны , расположенные в передних рогах спинного мозга , несколько дорсальнее и медиальнее, чем мотонейроны (МН). Это небольшие клетки. Диаметр тела клетки равен 10-20 мкм, дендриты имеют длину в 100—150 мкм, аксоны этих клеток — длинные (до 12 мм).
Принцип работы
Одиночное мышечное сокращение длится довольно долго (много десятков миллисекунд). Но следует учитывать то, что при напряжении мышцы, содержащей огромное количество , никогда не происходит одновременное их возбуждение . Активность различных мышечных волокон в какой-то степени чередуется, за счет этого мышца меньше утомляется. Поэтому для поддержания непрерывного мышечного напряжения не нужна высокая частота разряда двигательной нервной клетки . Для этого достаточна частота импульсации, не превышающая десяти импульсов в секунду. Мотонейроны имеют механизмы, которые стабилизируют их разряд именно на такой частоте и предотвращают возникновение импульсации слишком высокой частоты, которая могла бы привести к нарушению мышечной деятельности. Таким стабилизирующим механизмом является, во-первых, развитие в соме мотонейрона длительной следовой гиперполяризации после генерации импульса. Длительность её достигает примерно 100 мс, и в период её развития новое синаптическое действие будет ослаблено. Этот механизм сам по себе должен способствовать стабилизации частоты разряда мотонейрона на уровне около 10 импульсов в секунду. Кроме внутреннего механизма стабилизации, у мотонейрона есть еще и второй, внешний механизм, который работает в том же направлении. Этот внешний механизм представлен короткой цепочкой отрицательной обратной связи , через которую мотонейрон сам себя тормозит, но в том случае, когда он посылает разряд в аксон.
Общая схема деятельности такой цепочки выглядит следующим образом. На клетках Реншоу заканчиваются возвратные коллатерали аксонов, которые в пределах серого вещества отдают альфа-мотонейроны , иннервирующие двигательные мышцы, и поэтому они всегда «знают», насколько сильно возбужден нейрон. Клетки Реншоу, в свою очередь, заканчиваются на мотонейронах тормозными синапсами . Следовая гиперполяризация в клетках Реншоу отсутствует, и поэтому они могут на одном синаптическом потенциале генерировать целую пачку импульсов с очень высокой частотой — до 1500 импульсов в секунду. Каждый из этих импульсов, приходя к мотонейронам, вызывает в них тормозящую реакцию, которая суммируется до тех пор, пока длится разряд клетки Реншоу. Поэтому общая длительность торможения после одиночного импульса в аксонной коллатерали достигает примерно 100 мс. Возвратное торможение складывается со следовой гиперполяризацией и еще больше способствует удерживанию разряда мотонейрона на низкой частоте. Клетки Реншоу получают сигнал более, чем от одного мотонейрона, и сами посылают аксоны к многим мотонейронам. Поскольку возникли такие эффективные дублирующие друг друга механизмы стабилизации разряда мотонейрона, то очевидно, что последний из них имеет существенное значение для нормального осуществления двигательного акта.
В качестве нейромедиатора клетки Реншоу используют глицин — тормозной медиатор, действующий на альфа-мотонейроны.
Частота импульсов, посылаемых клеткой Реншоу, в широком диапазоне прямо пропорциональна частоте импульсов, посылаемых связанным с ней мотонейроном, а частота импульсов мотонейрона обратно пропорциональна частоте импульсов, посылаемого клеткой Реншоу. Клетки Реншоу играют роль «ограничителей» или «регуляторов» системы альфа-мотонейронов, и, таким образом, помогают предотвратить тетанус и повреждение мышцы. Благодаря их активности импульсация мотонейронов поддерживается в оптимальном диапазоне, необходимом для управляемого сокращения мышц.
Рефлекторная функция
Английский физиолог Шеррингтон впервые показал, что в любом рефлекторном акте участвуют процессы как возбуждения, так и торможения. При сокращении какой-то группы скелетных мышц тормозятся центры . При сгибании руки или ноги центры мышц-разгибателей затормаживаются. Рефлекторный акт возможен только при сопряженном, так называемом реципрокном торможении мышц-антагонистов. При ходьбе сгибание ноги сопровождается расслаблением мышц-разгибателей и, наоборот, при разгибании тормозятся мышцы-сгибатели. Если бы этого не происходило, то возникла бы механическая борьба мышц, судороги.
При раздражении чувствительного нерва, вызывающего, например, сгибательный рефлекс , импульсы направляются через спинной мозг к центрам мышц-сгибателей и к центрам мышц-разгибателей. В первых они вызывают процесс возбуждения, а во вторых с помощью клеток Реншоу — торможения. В результате возникает координированный, согласованный рефлекторный акт — сгибательный рефлекс .
Функции
- Одна из самых важных функций клеток Реншоу, общих для всех типов мотонейронов , — это стабилизация частоты их работы при поддержании позы или удержании нагрузки. То есть КР сглаживают колебания на выходе МН. Когда входной поток к МН возрастает, МН импульсируют сильнее, частота работы КР увеличивается и они притормаживают МН.
Эта функция была предложена Гессе и др. в 1975 году. Эта идея очень естественна, ведь МН с КР — типичная система с отрицательной обратной связью. В 70-х годах XX века такие системы усиленно обсуждались кибернетиками.
- КР играют важную роль в управлении МН-пулом, состоящем из МН разных типов. Например, когда человек постепенно напрягает мышцы руки, по принципу величины сначала включаются медленные моторные единицы, потом частота импульсов нарастает до 50 Гц, потом включается в работу FR (fatigue-resistant; fast) МН, но при этом частота S-МНа (slow) не растет. Раз включились FR-МН, значит приток импульсов к данному МН пулу увеличился. Частота S-МН не возрастает, так как FR-МН возбуждают КР, которые стабилизируют частоту S-МН. Это необходимо для защиты нейронов от «перегрузки», то есть чтобы нейрон не работал долго с высокой частотой, так как они от этого умирают.
Таким образом, еще одна функция КР — защита медленных МН от гибели. Высокая чувствительность S-МН к возвратному торможению была показана Гранитом в 1957 г. А в 1960 г. он высказал соображения о том, что возвратное торможение стабилизирует частоту разряда S-МН. Пока работали только S-МН, то есть частота не превышала 50 Гц, они не активировали сколько-нибудь заметного числа КР, и с ростом притока импульсов возрастала частота этих МН.
- В 1971 г. Халтборн и Янковская показали, что КР тормозят интернейрон реципрокного (взаимного) торможения.
Даже при очень сильном возбуждении МН синергиста (мышцы) сильно затормозить МН антагониста невозможно. Чем выше частота импульсации по волокнам 1а, тем сильнее должно быть торможение МН антагониста, но при этом сильнее работают МН синергиста, а значит и возбуждаемые ими клетки Реншоу. КР тормозят тормозной интернейрон, чтобы мышца-антагонист не оказалась сильно заторможенной и смогла бы быстро среагировать на возбуждающий импульс. Это нужно, чтобы поочередность работы мышц-антагонистов соблюдалась, их реакция была бы быстрой.
Когда начинает работать мышца-антагонист аналогичное реципрокное торможение осуществляется для МН мышцы-синергиста.
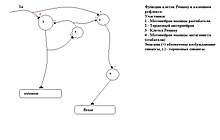
- Еще одна функция КР — десинхронизация МН одного пула. Её механизм плохо изучен.
- Одна из дополнительных функций КР состоит в том, что в быстрых, резких движениях, например, отряхивательном рефлексе, медленные МЕ могут помешать его осуществлению, для этого их деятельность надо подавить, то есть их затормозить.
- Еще одна функция КР. Фиффе (1991) показал, что КР образуют синапсы на дендритах МН, в отличие от реципрокного торможения, когда они образуют синапсы на телах МН. Эти синапсы позволяют избирательно тормозить некоторые входы в клетку, без торможения остальных входов в МН.
Все перечисленные выше функции КР были внутри одного МН пула, но есть ряд гипотез о их функциях на уровне координации работы пулов разных мышц.
- КР дают коллатерали своих аксонов к МН мышц-синергистов одного сустава и к МН флексоров (или экстензоров) разных суставов конечности. Тем самым КР связывают МН пулы разных мышц в единый МН пул.
- Еще одна идея состоит в том, что моторные команды, приходящие из коры полушарий головного мозга могут возбуждать нужные для движения МН и КР, которые тормозят МН мышц, которые не должны быть задействованы в этом движении. Таким образом, предполагается, что КР — часть механизма управления произвольными движениями!
Существует еще несколько функций клеток Реншоу. Например, Рилл в 1970 году показал, что КР могут тормозить другие КР, при этом КР агонистов сильнее тормозят КР антагонистов. КР могут тормозить нейроны вентрального спиномозжечкового тракта и другие нейроны восходящих трактов. Большинство описанных результатов было получено на МН задней лапы кошки. Оказалось, что эффективность возвратного торможения в других суставах иная.
Нарушения
Существует ряд веществ, которые подавляют активность клеток Реншоу. Наиболее известные из них — стрихнин и токсин Clostridium tetani (возбудитель столбняка ).
Стрихнин специфически действует на способность клеток Реншоу контролировать работу альфа-мотонейронов. Он является антагонистом нейромедиатора глицина и блокирует его рецепторы на альфа-мотонейронах и других нейронах. В результате торможения альфа-мотонейронов не происходит, поэтому возникают неконтролируемые сокращения мышц (судороги). Стрихнин может вызвать летальный исход из-за его воздействия на дыхательные мышцы, в том числе диафрагму , так как блокируется возможность совершать дыхательные движения.
Клетки Реншоу также являются мишенью для токсина Clostridium tetani , спорообразующей анаэробной бактерии, живущей в почве. Когда С.tetani попадает в организм через повреждения кожных покровов и её токсин с током крови попадает в спинной мозг, происходит нарушение выделения глицина и блокируется передача тормозного влияния с клеток Реншоу на альфа-мотонейроны. В результате альфа-мотонейроны становятся гиперактивными, и мышцы начинают совершать тетанические сокращения. Судороги охватывают большие группы мышц и в тяжелых случаях могут продолжаться почти непрерывно. Летальный исход может наступить на высоте судорог от асфиксии вследствие спазма мышц гортани в сочетании с уменьшением легочной вентиляции из-за напряжения межреберных мышц и диафрагмы. Кроме того, причиной смерти может стать непосредственное поражение дыхательного и сосудисто-двигательного центров ствола мозга.
Литература
- В. В. Шульговский «Основы нейрофизиологии» учебное пособие для студентов вузов 2-е издание М.АспектПресс, 2005.
- «Физиология человека» под редакцией В. М. Покровского, Г. Ф. Коротько 2-е издание изд. «Медицина»
- Савельев А.В. Моделирование систем клеток Реншоу // В сб.: «Моделирование неравновесных систем». Красноярск: ИВМ СО РАН, 2003. С. 143-145.
См. также
|
Нейроны
( Серое вещество ) |
|
|---|---|
|
/
|
|
|
/
Моторный нейрон |
|
| Синапс | |
| Сенсорный рецептор | |
| Нейроглия | |
|
Миелин
( Белое вещество ) |
|
| Соединительная ткань |
|
- 2020-12-11
- 1

