Interested Article - Кабельная теория дендритов

- 2020-03-21
- 1
Кабельная теория дендритов (также Кабельная теория Ролла или Модель Ролла ) — это множество предположений и результатов, которые относятся к распространению и взаимодействия электрических сигналов в дендритных деревьях. Эта теория была разработана в 1959 году американским математиком и нейрофизиологом для объяснения влияния электротонических свойств и морфологии дендритов на нейроны .
История
Понимание электротонических свойств нейрона возникло в XIX в. как результат объединения теории распространения тока в нервных клетках и мышцах с кабельной теорией, которая описывала распространение электрического тока на дальних расстояниях через кабели на дне океана. Поэтому «электротонические свойства» нейронов часто называют «кабельными свойствами». Основы кабельной теории заложил английский физик Уильям Томсон (Лорд Кельвин) , который в 1855 представил Королевскому Обществу математический анализ затухания сигнала в трансатлантическом телефонном кабеле между Америкой и Англией, который тогда планировали прокладывать. Подводный кабель схож по своим свойствам с нервным волокном. Он имеет токопроводящие сердцевины, покрытые изолирующей оболочкой, и окружён морской водой . (Так же и нейроны находятся в соляном растворе, который содержит электрически заряженные частицы ( ионы ), которые генерируют электрический потенциал и двигаются в ответ на потенциалы). Но, поскольку изоляция кабеля не является совершенной, то существует конечное сопротивление утечки через изолятор. Главная количественная разница между кабелем и нейроном в том, что кабельные жилы сделаны из меди, что является намного лучшим проводником, чем соляной раствор нейрона, а также кабельное покрытие является значительно лучшим изолятором, чем мембрана клетки. Поэтому сигналы в кабеле, в отличие от нейрона, могут распространяться на дальние расстояния без существенного затухания .
Кабельная теория впервые была математически применена к нервной системе в конце XIX в. для описания электрического тока через нервные волокна. В 1940-х гг. Алан Ходжкин и др. применили кабельную теорию к аксонам беспозвоночных ( краба и кальмара ), тем самым положив начало математическому описанию и моделированию нейронов. Однако применение кабельной теории к дендритам считалось математически невозможным в связи с их сложной разветвлённой структурой . Лишь в 1960-х и 1970-х гг. математик и нейробиолог Вилфрид Ролл из Национальных институтов здоровья в серии статей , показал, как эта проблема может быть решена с помощью использования метода вычислительного компартментного моделирования ( англ. compartmental modeling ), и он же впервые применил кабельную теорию для описания интегративных функций дендритов, заложив основы вычислительной нейронауки . В те времена считалось, что дендриты являются настолько электротонически длинными, что любые входящие импульсы к ним не могут существенно влиять на электрические свойства сомы . Однако, соединив математическую кабельную теорию и те незначительные физиологические данные, которые в то время существовали, Ролл убедительно показал, что классические представления о дендритах существенно недооценивают их значительный вклад в нейронные исчисления и передачу информации . Созданные Роллом модели позволили построить единую математическую теорию дендритных функций, которая в сочетании с математическими моделями генерации синаптических потенциалов и потенциала действия обеспечила целостное теоретическое описание нейронной активности.
Кабельная теория Ролла
Основные предположения теории
Фундаментальным законом, описывающим распространение электротонического тока по дендриту, является простое уравнение закона Ома :

Математическое описание прохождения тока по дендритам основывается на нескольких базовых предположениях:
- Мембрана в кабельной теории является пассивной (независима от напряжения) и однородной, тогда как реальные дендриты коры обладают нелинейными мембранными каналами.
- Входами к кабелю являются токи, которые линейно суммируются, в отличие от изменений в синаптической проводимости мембраны, которые не суммируются линейно.
- Электротонический ток испытывает влияние двух различных сопротивлений : внутреннего (или осевого) и мембранного сопротивления. Для простоты описания пассивного распространения потенциала, при установившемся входе к кабелю или дендриту с однородным потенциалом покоя ( E r ), E r и ёмкость C m могут быть проигнорированы. Следовательно, ток только делится на каждой точке прохождения через две линии сопротивления: осевого и мембранного.
- Поскольку, как известно, придание параллельных сопротивлений уменьшает общее сопротивление, то осевое сопротивление является обратно пропорциональным диаметру кабеля или дендрита,

где r i — осевое сопротивление на единицу длины (в Ω·см осевой длины) и А — площадь поперечного сечения ( π r 2 ). Поэтому толстый цилиндр или дендрит имеет меньшее осевое сопротивление, чем тонкий. Следовательно, предполагая, что осевое сопротивление является однородным по всему кабелю или дендриту, оно может быть выражено как

где R i — удельное осевое или внутреннее сопротивление (Ω·см осевой длины). Это является одним из ключевых положений кабельной теории. Дендриты являются тонкими трубочками, окутанными мембраной, которая является относительно хорошим электрическим изолятором по сопротивлению от внутриклеточной сердцевины или внеклеточной жидкости, поэтому изменения потенциала в радиальных или осевых направлениях являются незначительными в сравнении с продольными изменениями. Благодаря этой разнице в мембранном и осевом сопротивлении, для короткой длины дендрита электрический ток внутри серцивины кабеля (core conductor) проходит параллельно оси цилиндра (вдоль оси x ). Именно поэтому классическая кабельная теория рассматривает только одно пространственное измерение ( x ) вдоль кабеля, игнорируя y и z измерения. Другими словами, ключевое предположение одномерной кабельной теории заключается в том, что мембранный потенциал вдоль нейронного кабеля выражается как функция одной продольной пространственной координаты x и времени t, t, V(x, t) .
Следует, конечно, иметь в виду, что реальные дендриты — это не пустые цилиндры, и они являются трёхмерными. Наличие цитоскелетных элементов, органелл увеличивает эффективное внутреннее сопротивление, или осевое сопротивление, тем самым влияя на распространение электротонических сигналов.
Эквивалентный цилиндр
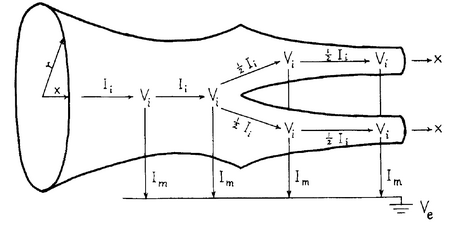
Кабельная модель дендритов (кабельную модель нейрона см., напр., в или более подробно в , , ) математически значительно сложнее, чем аксонов, поскольку их относительно малая длина является ключевым фактором для построения электротонических свойств. Для того, чтобы преодолеть эту сложность, линейная кабельная теория трактует каждый сегмент дендрита как кабель или цилиндр, окружённый пассивной мембраной, и который имеет постоянный радиус, по которому распространяется электротонический потенциал, что позволяет моделировать его через омическую проводимость, параллельную с ёмкостью. Поскольку внутри дендритного дерева нет петель, и дендритные ветви не связаны между собой непосредственно, то они могут рассматриваться как настоящие деревья в смысле теории графов , с единичным путём между любыми двумя точками на дереве. Прохождение тока в таких деревьях описывается линейным одномерным пассивным кабельным уравнением ( дифференциальное уравнение в частных производных )

где V — разница потенциалов вдоль мембраны, — это пространственная постоянная (постоянная длины) [1] нервного волокна, которая выражает расстояние, после прохождения которого потенциал уменьшается до 1/ e своей исходной амплитуды (то есть, до 37 % от первоначального значения); r m — мембранное сопротивление (в Ω·см); — это мембранная временная постоянная, а c m (в Ф/см) — мембранная ёмкость. Математическая сложность заключалась в решении этого уравнения для произвольных дендритных структур. Найденное Роллом аналитическое решение позволило ему показать существенное влияние дендритов на электрическое поведение нейронов.


Кабельное уравнение конечного кабеля является справедливым для любого неразветвлённого цилиндрического сегмента с постоянными мембранными свойствами и постоянным диаметром. Однако остаётся вопрос, можно ли использовать это уравнение для изучения потенциала в двух или более соединенных вместе сегментах. Поскольку то, каким образом сегменты необходимого кабеля соединены, зависит от надлежащего выбора граничных условий для каждого сегмента, то предположим, пусть три ветви имеют электротонические длины L 0 , L 1 и L 2 , с бесконечными входными сопротивлениями R ∞.0 , R ∞.1 и R ∞.2 . Как известно из кабельной теории, бесконечное входное сопротивление конечного кабеля равно входному сопротивлению полубесконечного кабеля такого же диаметра и имеет такие же значения R m и R i , и вычисляется в соответствии с
- (1)

Для простоты предположим, что две самые крайние ветви заканчиваются запечатанными концами (граничные условия), то есть осевой ток не проходит в кабель. Входные сопротивления двух самых крайних дочерних ветвей в точке разветвления, глядя в сторону их концов, задаются через уравнения входного сопротивления для бесконечного кабеля как

- (2)

и
- (3)

Правая сторона главного кабеля заканчивается в точке разветвления с двумя дочерними кабелями. Его нагрузочное сопротивление определяется входными сопротивлениями дочерних ветвей, в силу того факта, что полная проводимость двух параллельных проводимостей равна их сумме. Иначе говоря, мы вычисляем нагрузочное сопротивление, эффективно заменяя две проводимости дочерних ветвей одной. Тогда эффективное нагрузочное сопротивление главного кабеля определяется через

- (4)

Наконец, используя кабельные уравнения для напряжения и входного сопротивления в конечном кабеле, входное сопротивление на левом конце главной ветви (глядя в сторону дочерних ветвей) определяется через
- (5)

Этот алгоритм может быть рекурсивно применён для расчёта входного сопротивления произвольных пассивных деревьев и таким образом может позволить точно вычислить входное сопротивление на любой точке дерева, в чём и заключалось одно из важных достижений Ролла.
После расчёта входного сопротивления в любом конкретном месте на дендритном дереве, мы можем вывести устоявшееся напряжение в данном месте в ответ на поданный ток , используя закон Ома . К примеру, напряжение в точке x = 0 в родительском кабеле будет

- (6)

С учётом V 0 и используя кабельное уравнение для напряжений в конечном кабеле, мы можем вычислить изменение напряжения везде вдоль главной ветви в ответ на подачу установившегося тока I inj . Тогда, если, например, родительский кабель поставляет ток I 0 двум дочерним ветвям, и нам нужно узнать, какая часть I 1 этого тока будет идти в один конец, а какая часть I 2 — в другой, то используя уравнение закона Ома для дискретного кабеля, входное сопротивление в полубесконечном кабеле и неймановские граничные условия, мы можем вычислить ток в одном дочернем кабеле (предполагая, что его конец запечатан, хотя это и не обязательно),






- (7)

где V 1 — напряжение в точке ветвления. В этой точке, X = 0 (для дочерней точки), и ток, протекающий в этой ветке, определяется через
- (8)

Поскольку тот же принцип применяется ко второй дочерней ветви и является справедливым для произвольного граничного условия, то отсюда следует, что
- (9)

где и так же для . Другими словами, ток распределяется между этими ветвями в соответствии с их входными проводимостями.


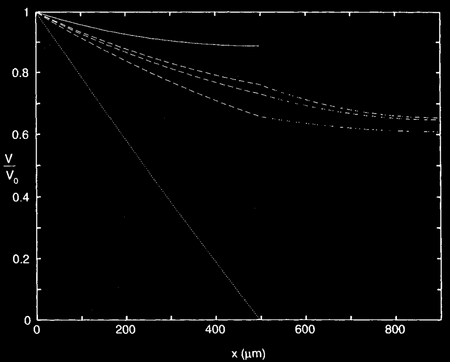
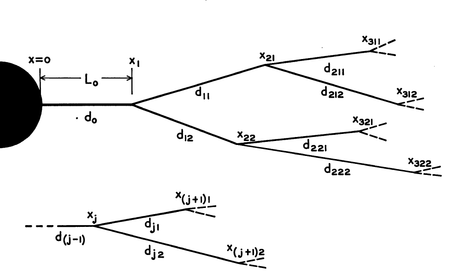
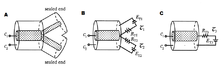
Позже Ролл нашёл ещё более простой и изысканный способ вывода потенциала в разветвлённом дереве, который стал вершиной его кабельной теории дендритов Вычислим распределение напряжений в простом дереве (см. рисунок), предполагая, что две дочерние ветви являются идентичными (то есть L 1 = L 2 и d 1 = d 2 с одинаковыми параметрами мембраны) и что установившийся ток подаётся в левый конец родительского цилиндра. Нормированный уровень напряжения через это минидерево показан на рисунке для трех различных комбинаций диаметров родительских и дочерних ветвей: (1) , (2) і (3) Верхняя и нижняя кривые показывают два предельных случая, в которых родительский кабель ограничен с правого конца, или с «граничными условиями для запечатанного конца», или в коротком замыкании. В случаях же, когда родительский кабель в диаметре больше вдвое дочерних ветвей, или имеет тот же размер, то, как видно на рисунке, будет разрыв в производной напряжения в точке ветвления.



В третьем случае, , все производные уровня напряжения являются непрерывными, и происходит интересная вещь: в отличие от двух других случаев, падение напряжения вдоль обеих линий кабелей может быть описано одним выражением. Для объяснения этого феномена воспользуемся кабельным уравнением, описывающим напряжение в конечном кабеле длины L 0 с произвольными начальными условиями, определёнными через нагрузочное сопротивление R L.0 :

- (10)

Поскольку оба дочерних кабеля идентичны и, следовательно, имеют идентичное входное сопротивление, (уравн. 2 и 3), то из уравнения 4 следует, что объединённые входные сопротивления обоих кабелей будут

- (11)

Возвращаясь к уравнению 1 для входного сопротивления полубесконечного кабеля, можно отметить обратную зависимость диаметра кабеля в степени 3/2:
- (12)

Если тогда

- (13)

То есть, бесконечное входное сопротивление родительского кабеля согласуется с бесконечным входным сопротивлением двух параллельных дочерних ветвей. Сочетая уравн. 11 и 13, получаем
- (14)

Помещая это обратно в уравнение 10, получаем
- (15)


Это уравнение описывает напряжение в одном, неразветвлённом (без ветвей) цилиндре с длиной с граничными условиями для запечатанного конца. Итак, вместо того, чтобы моделировать эту структуру как три взаимосвязанные цилиндра, можно свести их к единому «эквивалентному цилиндру», с электротонической длиной и диаметром, одинаковым с диаметром главной ветви. Главной причиной, почему такое упрощение возможно, является то, что бесконечное входное сопротивление главной ветви «согласуется» с бесконечным входным сопротивлением двух дочерних параллельных ветвей, — то, что в электронике называется «согласование импедансов». В частности, напряжение и входное сопротивление теперь имеют непрерывные производные вдоль точки ветвления (см. рисунок). В целом же, согласование импедансов не означает, что входное сопротивление R in дочерних кабелей согласовано с главным кабелем (поскольку входное сопротивление главного кабеля в правом конце пропорционально cth L 0 , тогда входное сопротивление дочерних ветвей пропорционально cth L 1 ).


В. Ролл показал, что весь класс дендритных деревьев может быть сведён к единому эквивалентному цилиндру, если только выполняются четыре главных условия:
- Значение R m и R i одинаковы во всех ветвях;
- Граничные условия одинаковы на всех концах кабелей;
- Все конечные ветви заканчиваются на том же электротоническом расстоянии L от начала главной ветки, где L — это сумма значений L i от начала до удаленного конца каждой ветви; L соответствует общей электротонической длине эквивалентного цилиндра.
- В каждой точке разветвления бесконечные входные сопротивления должны согласовываться.
Если все кабели имеют то же мембранное сопротивление и внутриклеточную сопротивляемость, то это означает, что

Последнее выражение известно как закон .

Если эти четыре условия выполнены, то эквивалентный цилиндр прекрасно имитирует поведение всего дендритного дерева. Другими словами, инжекция тока в левый конец дерева даст такую же чувствительность напряжения, как и инжекция идентичного тока его эквивалентный цилиндр.
Для сложных дендритных структур, начиная с концов, можно рекурсивно упростить модель до одного полубескуонечного цилиндра. До этого рассматривался лишь пример с двумя ветвями в точке разветвления, но мы можем иметь любое количество веток в каждой точке разветвления, пока

где d P — диаметр родительского дендрита, а d D — диаметры дочерних дендритов. Если это условие сохраняется в каждой точке разветвления и материальные свойства кабелей те же самые, тогда всё дендритное дерево может быть сведено к эквивалентному полубескуонечному кабелю. С таким эквивалентным цилиндром вся сложность дерева выражается четырьмя главными биофизическими параметрами:
- кабельной длиной L дендритного дерева ( λ ),
- мембранной временной постоянной ( τ m ),
- входным сопротивлением в конце сомы ( R in ),
- отношением между входной проводимостью дендритов и сомы ( p ).
Сведя сложную морфологию дендритов к «эквивалентному цилиндру», кабельная теория Вилфрида Ролла позволила описать электрические и вычислительные свойства пассивных дендритных деревьев с любой сложностью, в частности аналитически вычислить чувствительность напряжения в произвольной точке дендритного дерева к току, поданному в любую другую точку. Это дало возможность оценить амплитуду синаптического потенциала и его зависимость от времени в данном дендритном месте, как и обнаружить, что кабельные свойства дендритов (быстрая зарядка их мембранной ёмкости) фильтруют высокие временные частоты, которые составляют постсинаптические потенциалы (ПСП), тем самым позволяя дендритам действовать как фильтр низких частот . Кроме этого, определённый процент синаптического тока вытекает через дендритную мембрану и, как следствие, ПСП перетерпевают значительное затухание напряжения и временную задержку, пока они распространяются к аксону, чтобы вызвать потенциал действия .
Ролл впервые аналитически показал, что спад напряжения в распределённых (не-изопотенциальных) пассивных дендритных деревьях определяется суммой многих экспоненциальных спадов, а не одной экспонентой , как в точечной RC -модели : чем дальше входной импульс от сомы , тем медленнее время нарастания и шире конечный соматический ПСП. Для быстрых ПСП, как следовало из модели, пик затухания должен быть стократным и существенно замедлиться (от 5 до 50 мсек), уходя от удалённого дендрита к соме. Как позже стало ясно, подобная временная задержка дендритных ПСП позволяет нейронам вычислять направление движения . В отличие от существенного затухания пиков ПСП, значительная доля синаптического заряда (50 % для удаленных синапсов) достигает сомы. Это связано с тем, что внутриклеточное (осевое) сопротивление дендритов является значительно меньшим, чем мембранное сопротивление, поэтому только относительно небольшой процент синаптического заряда теряется через дендритное мембранное сопротивление. Это имеет важное следствие для степени локальной взаимосвязи между синапсами в дендритном дереве (т. наз. уровень электрического сегментирования), как и для обратного распространения потенциала действия от сомы к дендритам и, с другой стороны, от дендритов к соме. Таким образом, даже в пассивных дендритах отдалённые синапсы способны существенно влиять на выходной импульс аксона, способствуя значительной деполяризации сомы.
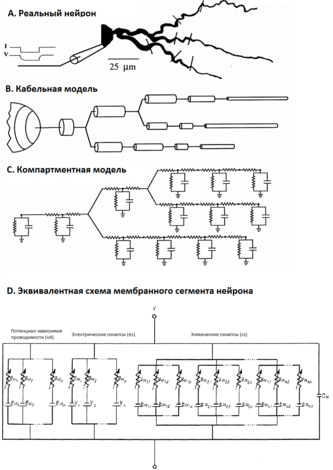
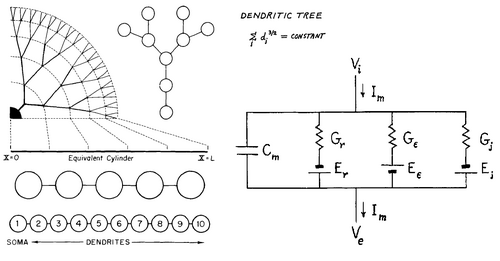
A . Частичная геометрия реального нейрона с сомой и дендритными разветвлениями.
B. Кабельная модель: нейрон как множество цилиндрических кабелей. Напряжение в любой точке дендритного дерева вычисляется с помощью кабельного уравнения и необходимых граничных условий (см. текст).
C . Компартментная модель. Дендритное дерево дискретизируется на множество взаимосвязанных RC -сегментов , каждый из которых является моделью с сосредоточенными параметрами для представления дендритных частей. Мембранные сегменты соединены через осевое, цитоплазматическое сопротивление. Здесь напряжение может быть вычислено на каждом сегменте для любого нелинейного входного сигнала и мембранных свойств, зависящих от времени и напряжения.
D. Электрическая эквивалентная схема мембранного сегмента нейрона, с мембранным потенциалом V и ёмкостью C m .
Другой важный вывод, который вытекает из кабельной теории дендритов, заключался в том, что дендриты накладывают на сому огромную проводимость нагрузки и, как следствие, значительная часть тока, подаваемого через электрод в сому, идёт в дендритное дерево. Результатом этого является усиление частоты зарядки/разрядки соматической мембраны по сравнению с сомой без дендритов. Также кабельная теория объяснила электрические последствия наличия сложных геометрий дендритных деревьев . В частности, на отдалённых дендритных ветвях и шипиках должен существовать большой входной импеданс — и как следствие, большое локальное изменение напряжения, — в несколько ГОм. Одно из наиболее важных применений кабельной теории связано с оценкой биофизических параметров реальных нейронов, — основываясь на экспериментальных данных, — особенно мембранного и цитоплазматического сопротивлений ( R m и R i ), и электротонической длины L .
Кабельная теория, разработанная Роллом, продемонстрировала, что комбинация специфической морфологии и электрических свойств мембраны и цитоплазмы (в частности, электротонической структуры) определяет обработку и распространение электрических сигналов в дендритах. Иначе говоря, даже два морфологически идентичных дендритных дерева, но с различными электрическими свойствами, могут иметь совершенно разные вычислительные характеристики.
Компартментные модели
При всей их значительной концептуальной привлекательности, аналитические решения кабельного уравнения становятся всё более громоздкими, когда мы имеем дело с активными, развитыми и неоднородными дендритами, в которых вместо источники токов — реальные синапсы , которые меняют электрические свойства постсинаптической мембраны. Один из способов решения этой проблемы был предложен В. Роллом — это т. наз. «компартментное моделирование» ( англ. compartmental modeling ), которое он позаимствовал из химической инженерии и которое представляет собой математически конечно-разностную (дискретную) аппроксимацию линейного кабельного уравнения или его нелинейных расширений.
Этот метод заменяет непрерывное кабельное уравнение множеством, или матрицей , обыкновенных дифференциальных уравнений (которых иногда может быть до тысячи), для решения которых используют вычислительные методы для каждого интервала времени. В компартментной модели дендритные сегменты, которые являются электрически короткими, рассматриваются изопотенциальными (электрически однородными) цилиндрами, и сосредоточены в единый RC (или пассивный, или активный) мембранный компартмент (отсек). Компартменты соединены между собой через продольное сопротивление согласно топологии дерева. Тогда различия в физических свойствах (диаметр, мембранные параметры) и разнице потенциалов возникают уже между компартментами, а не в них самих. Этот подход позволил Роллу описать дендриты как серии анатомических сегментов, мембранный потенциал которых является функцией от тока, протекающего в или из сегмента.
Сила и производительность компартментного моделирования быстро стала очевидной, поскольку оно было достаточно гибким и позволяло включать в модели параметры с разным уровнем функциональной сложности. Пока данный объект изучения ( ионный канал , мембранная проводимость и др.) может быть описан дифференциальным или стационарным уравнением, он может быть включен в компартментную модель, давая возможность с помощью таких программных пакетов, как NEURON или GENESIS симулировать его свойства на основе экспериментальных данных .
Использование вычислительных методов для определения изменения мембранного потенциала вдоль дендритного дерева позволило поместить в компартменты нелинейные ионные каналы, тем самым делая возможным изучение нелинейной интеграции синаптических входных импульсов в дендритах. Одним из последствий изменения проводимости, связанной с синаптическим импульсом, является то, что синапсы нелинейно взаимодействуют друг с другом. Компартментные модели с пассивными дендритами позволили показать, что прилегающие дендритные синапсы суммируются менее линейно друг с другом, чем отдалённые синапсы. Подобная чувствительность к пространственной организации синапсов означала, что локальные нелинейные синаптические операции могут выполняться полунезависимо во многих дендритных субъединицах .
Другая важная черта дендритных синапсов, впервые обнаруженная с помощью компартментного моделирования, заключается в том, что они способны эффективно масштабировать кабельные свойства дендритного дерева. Когда тысячи синапсов поступают к дереву, дендритная мембрана становится сильно «протекающей» и, как следствие, кабельные свойства дендритов существенно меняются: R in и τ m убывают с активностью, тогда как L растёт. Так же кардинально меняется временное разрешение (чувствительность к синхронизации входных импульсов, которая зависит от эффективного τ m ) и временная замедленность дендритов, как результат синаптической активности .
Впрочем, наряду с эффективностью компартментных методов, возникают значительные практические (увеличение сложности дифференциальных уравнений, лежащих в их основе) и концептуальные проблемы (увеличение количества свободных параметров). Тем более, пока нет экспериментальных и количественных данных по распределению специфического типа ионных каналов вдоль дендритного дерева, что бы позволяло строить приближенные к реальным нейронам модели. Чтобы избежать этих трудностей, все нелинейные ионные каналы, которые отвечают за генерацию спайков, обычно помещаются вместе на соме, и дендритное дерево трактуется как пассивный кабель, что значительно упрощает его сложное поведение .
Примечания
- ↑ The Theoretical Foundation of Dendritic Function: Selected Papers of Wilfrid Rall with Commentaries (англ.) / Segev I., Rinzel J. and Shepherd G.. — MIT Press , 1995. — P. 456. — ISBN 9780262193566 .
- Koch, Christof. Biophysics of Computation: Information Processing in Single Neurons (англ.) . — Oxford University Press , 2004. — P. 562. — ISBN 9780195181999 .
- Korogod Sergey M. and Suzanne Tyč-Dumont. Electrical Dynamics of the Dendritic Space (англ.) . — Cambridge University Press , 2009. — P. 211. — ISBN 9781139481229 .
- ↑ Rall Wilfrid. (англ.) // : journal. — 1959. — November ( vol. 1 , no. 5 ). — P. 491—527 . — doi : . 4 ноября 2012 года.
- ↑ Rinzel John and Rall Wilfrid. (англ.) // : journal. — 1974. — October ( vol. 14 , no. 10 ). — P. 759—790 . — doi : . 24 сентября 2015 года.
- ↑ Rall Wilfrid. (англ.) // : journal. — Vol. 96 . — P. 1071—1092 . — doi : . 13 октября 2013 года.
- ↑ Rall Wilfrid. (англ.) // : journal. — Vol. 2 . — P. 145—167 . 15 октября 2022 года.
- ↑ Rall Wilfrid and Rinzel John. (англ.) // : journal. — 1973. — July ( vol. 13 ). — P. 648—688 . — doi : . 24 сентября 2015 года.
- Rall Wilfrid. (англ.) // : journal. — 1969. — December ( vol. 9 ). — P. 1483—1508 . — doi : . 24 сентября 2015 года.
- Segev Idan. (англ.) // : journal. — 1992. — November ( vol. 15 ). — P. 414—421 . — doi : . 11 марта 2012 года.
- Coombs J. S. ,Eccles J. C. , and Fatt P. (англ.) // : journal. — 1955. — 2 November ( vol. 130 ). — P. 374—395 .
- Николлс, Джон Г. От нейрона к мозгу (неопр.) . — 2012. — С. 671. — ISBN 9785397022163 .
- Jack James , Noble Denis , Tsien Richard W. Electric Current Flow in Excitable Cells (англ.) . — Oxford University Press , 1975. — P. 518.
- Rall, Wilfrid. Core Conductor Theory and Cable Properties of Neurons // Handbook of Physiology, The Nervous System, Cellular Biology of Neurons (англ.) / Mountcastle, Vernon B.. — , 1977. — P. 39—97. — ISBN 0683045059 .
- Tuckwell, Henry C. Introduction to Theoretical Neurobiology: Volume 1, Linear Cable Theory and Dendritic Structure (англ.) . — Cambridge University Press , 1988. — P. 304. — ISBN 9780521350969 .
- ↑ Rall, Wilfrid. Theoretical significance of dendritic trees for neuronal input-output relations. // Neural Theory and Modeling: Proceedings of the 1962 Ojai Symposium (англ.) . — Stanford University Press , 1964. — P. 73—97. — ISBN 9780804701945 .
- Koch C., Poggio T. and Torres V. Retinal Ganglion Cells: A Functional Interpretation of Dendritic Morphology (англ.) // Philosophical Transactions of the Royal Society B: Biological Sciences : journal. — 1982. — Vol. 298 , no. 1090 . — P. 227—263 . — ISSN . — doi : .
- Rall W., Burke R.E., Holmes W.R., Jack J.J., Redman S.J. and Segev I. (англ.) // : journal. — 1992. — 1 October ( vol. 72 ). — P. 159—186 . 7 марта 2016 года.
- Vermeulen Arthur and Rospars Jean-Pierre. A simple analytical method for determining the steady-state potential in models of geometrically complex neurons (англ.) // : journal. — 1998. — Vol. 82 , no. 2 . — P. 123—133 . — ISSN . — doi : .
- Borg-Graham Lyle J. Additional Efficient Computation of Branched Nerve Equations: Adaptive Time Step and Ideal Voltage Clamp (англ.) // Journal of Computational Neuroscience : journal. — 2000. — Vol. 8 , no. 3 . — P. 209—226 . — ISSN . — doi : .
- Hines M. L. and Carnevale N. T. The NEURON Simulation Environment (англ.) // . — 1997. — Vol. 9 , no. 6 . — P. 1179—1209 . — ISSN . — doi : .
- Agmon-Snir Hagai, Carr Catherine E. and Rinzel John. The role of dendrites in auditory coincidence detection (англ.) // Nature : journal. — 1998. — Vol. 393 , no. 6682 . — P. 268—272 . — ISSN . — doi : .
- Segev I. and London M. Untangling Dendrites with Quantitative Models (англ.) // Science. — 2000. — Vol. 290 , no. 5492 . — P. 744—750 . — ISSN . — doi : .
- Gerstner Wulfram and Kistler Werner M. Spiking Neuron Models: Single Neurons, Populations, Plasticity (англ.) . — Cambridge University Press , 2002. — P. 480. — ISBN 9780521890793 .
Ссылки
- Кабельная теория — статья из Большой советской энциклопедии .
Литература
- Позин Н.В. Моделирование нейронных структур. М.: Наука, 1970.
- Позин Н.В.и др. Элементы теории биологических анализаторов. М.:Наука, 1978.
- Савельев А. В. Методология синаптической самоорганизации и проблема дистальных синапсов нейронов // Журнал проблем эволюции открытых систем . — Казахстан, Алматы, 2006. — Т. 8. — № 2. — С. 96-104.
- Николлс, Джон Г. От нейрона к мозгу (неопр.) . — 2012. — С. 671. — ISBN 9785397022163 .
- The Theoretical Foundation of Dendritic Function: Selected Papers of Wilfrid Rall with Commentaries (англ.) / Segev I., Rinzel J. and Shepherd G.. — MIT Press , 1995. — P. 456. — ISBN 9780262193566 .
- Tuckwell, Henry C. Introduction to Theoretical Neurobiology: Volume 1, Linear Cable Theory and Dendritic Structure (англ.) . — Cambridge University Press , 1988. — P. 304. — ISBN 9780521350969 .
- Koch, Christof. Biophysics of Computation: Information Processing in Single Neurons (англ.) . — Oxford University Press , 2004. — P. 562. — ISBN 9780195181999 .
- Shepherd, Gordon M. Creating Modern Neuroscience: The Revolutionary 1950s (англ.) . — Oxford University Press , 2009. — P. 304. — ISBN 9780199741472 .
- Ermentrout Bard and Terman David H. Mathematical Foundations of Neuroscience (неопр.) . — Springer, 2010. — С. 422. — ISBN 9780387877075 .
- 2020-03-21
- 1










