Горбацевич, Александр Алексеевич
- 1 year ago
- 0
- 0
Дендротоксины — класс пресинаптических нейротоксинов , вырабатываемых змеями рода мамба (и содержится в их яде ) и блокирующих определённые подтипы потенциалзависимых калиевых каналов в нейронах , тем самым повышая высвобождение ацетилхолина в нервно-мышечных синапсах . Благодаря высокой эффективности и селективности для калиевых каналов, дендротоксины оказались чрезвычайно полезными в качестве фармакологических средств для изучения структуры и функции этих белков ионных каналов .
Дендротоксины блокируют определенные подтипы потенциалзависимых каналов для калия (K + ) в нейронах. В нервной системе потенциалзависимые K + каналы отвечают за реполяризацию мембраны и контролируют длительность потенциалов действия . Дендротоксин, как было показано, связывается с калиевыми каналами перехватов Ранвье мотонейронов и блокирует активность калиевых каналов. Таким образом, дендротоксины увеличивают продолжительность потенциалов действия и повышают высвобождение ацетилхолина в нервно-мышечном синапсе, что может приводить к мышечной перевозбудимости и судорогам.
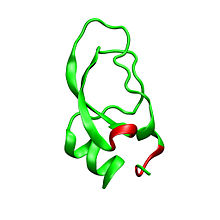
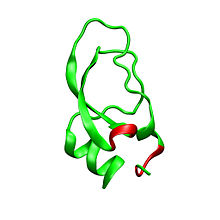
Дендротоксины - белки массой ~ 7kDa, состоящие из одной пептидной цепи из приблизительно 57-60 аминокислот. Были выделены несколько гомологов α-дендротоксина, все обладающие несколько иной последовательностью. Тем не менее, молекулярная архитектура и конформация этих белков очень похожи. Дендротоксины обладают очень короткой 3 10 -спиралью вблизи N-конца пептида, в то время как два оборота α-спирали происходит вблизи С-конца. Двунитевой антипараллельный β-лист занимает центральную часть молекулярной структуры. Эти две β-нити соединены искаженной областью β-оборота , что, предположительно, играет важную роль в активности связывания белка. Все дендротоксины сшиты тремя дисульфидными связями , которые добавляют стабильность белку и в значительной степени определяют его конформацию. Радикалы цистеина , образующие эти дисульфидные связи, идентичны по расположению у всех членов семейства дендротоксинов, они расположены в C7-C57, С16-С40, и С32-С53 участках (нумерация согласно α-дендротоксинам).
Дендротоксины структурно гомологичны ингибиторам сериновых протеаз (ИСП), в том числе апротинину . Было показано, что последовательности α-дендротоксина и ИСП идентичны на 35% и имеют идентичные дисульфидные связи. Несмотря на структурную гомологию между этими двумя белками, дендротоксины не оказывают никакого ощутимого ингибирующего действия на фактор свертывания, в отличие от ИСП. Эта потеря активности, возможно, является результатом отсутствия ключевых аминокислотных радикалов, которые производят структурные различия, препятствующие ключевым взаимодействиям, необходимым для активности ИСП.
Дендротоксины – основные белки, имеющие положительный заряд при нейтральном pH . Большинство положительно заряженных радикалов аминокислот находятся в нижней части структуры, создавая катионную область в одной части белка. Положительный заряд исходит от радикалов лизина (Lys) и аргинина (Arg), которые сконцентрированы на трёх основных участках: вблизи амино-конца (Arg3, Arg4, Lys5), вблизи C-конца (Arg54, Arg55) и узкой области β-оборота (Lys28, Lys29, Lys30) . Полагают, что эти положительно заряженные радикалы могут играть критичную роль в связывающей активности дендротоксинов, так как они могут взаимодействовать с анионными участками (отрицательно заряженными аминокислотами) в порах калиевых каналов.
Одна молекула дендротоксина обратимо связывается с калиевым каналом для того, чтобы оказать ингибирующий эффект. Предполагается что это взаимодействие инициируется электростатическими взаимодействиями между положительно заряженными радикалами аминокислот в катионной области дендротоксина и отрицательно заряженными радикалами в порах ионных каналов . Калиевые каналы, как и другие катионно-избирательные каналы, предположительно имеют облако отрицательных зарядов, находящихся перед входным отверстием в пору канала, которое помогает проводить ионы калия по пути проникновения. Обычно считается (но не доказано), что молекулы дендротоксина связываются с анионными связями вблизи внеклеточной поверхности канала и физически закрывают пору, тем самым предотвращая проведение ионов. Однако, Имреди и МакКиннон предположили, что дельта-дендротоксин может иметь отклонённое от центра место связывания на целевых белках, и может ингибировать канал путём изменения его структуры, а не физического блокирования пор.
Многие исследования были направлены на определение радикалов аминокислот, необходимых для связывающего действия дендротоксинов на целевые калиевые каналы. Харви и другие учёные использовали особые модификации, направленные на радикалы, для определения положительно заряженных радикалов, имеющих решающее значение для блокирующей активности l-дендротоксина. Они сообщили, что ацетилирование Lys5 вблизи области амино-конца и Lys29 вблизи области β-оборота, привело к существенному уменьшению сходства связывания l-дендротоксина. Похожие результаты были получены для K-дендротоксина при использовании мутагенеза для замены положительно заряженных остатков лизина и аргинина на нейтральные аланины . Эти результаты, наряду со многими другими, предположительно показали, что положительно заряженные радикалы лизина в половине амино-конца, в частности, Lys5 в спирали 3 10 , играют очень важную роль в связывании дендротоксина с целевыми калиевыми каналами. Остатки лизина в области β-оборота предоставили более смешанные результаты, по всей видимости, являясь биологически критическим в некоторых гомологах дендротоксина и не обязательными для других. Кроме того, мутация всей тройки лизина (К28-К29-К30) в Ala-Ala-Gly в α-дендротоксине привело к очень незначительным изменениям биологической активности.
Существует общее мнение, что сохраняющийся лизиновый радикал вблизи амино-конца (Lys5 в α-дендротоксине) имеет решающее значение для биологической активности всех дендротоксинов, в то время как дополнительные радикалы, такие как в области β-оборота могут играть роль в специфичности дендротоксина путем опосредования взаимодействий отдельных токсинов на их отдельные целевые области. Это не только помогает объяснить жёсткую специфичность некоторых дендротоксинов для различных подтипов потенциалозависимых каналов K + , но также объясняет различия в силе действия дендротоксинов на обычные каналы K + . Например, Ван и другие учёные показали, что взаимодействие K-дендротоксина с K V 1.1 обеспечивается с помощью его радикалов лизина как в амино-конце, так и в области β-оборота, в то время как α-дендротоксин-видимому, взаимодействует с его мишенью исключительно за счет амино-конца. Эта менее обширная взаимодействующая область может помочь объяснить, почему α-дендротоксин менее различающий в то время как K-дендротоксин строго избирателен по отношению к K V 1.1.
Калиевые каналы нейронов позвоночных имеют множество разновидностей, позволяющих им подстраивать свойства их электрических сигналов проявлением различных комбинаций подгрупп калиевых каналов. К тому же, так как они регулируют движения ионов через биологические мембраны, они являются важными во многих аспектах клеточной регуляции и передаче сигналов разных типов клеток. Следовательно, потенциалозависимые калиевые каналы являются целями для широкого спектра сильнодействующих биологических токсинов таких животных, как змеи, скорпионы , актинии и конических улиток . Так, очищение яда привело к выделению пептидных токсинов, таких, как дендротоксин, которые стали полезными фармакологическими инструментами для изучения калиевых каналов. В связи с их активностью и избирательностью к различным подтипам калиевых каналов, дендротоксины стали пригодными к использованию в качестве молекулярных зондов для структурного и функционального анализа этих белков. Это может помочь пониманию ролей, играемых отдельными типами каналов, а также помочь классифицированию разнообразных типов каналов . Кроме того, наличие меченых радиоактивными изотопами дендротоксинов предоставляет инструмент для изучения других источников в поисках новых токсинов, действующих на калиевые каналы, таких как класс , присутствующих в актиниях. И, наконец, структурная информация, представленная дендротоксином может дать ключ к синтезу лекарственных соединений , которые могут воздействовать на определенные классы калиевых каналов