Тирозинкиназа
- 1 year ago
- 0
- 0

| Рецепторные тирозинкиназы | |
|---|---|

|
|
| Идентификаторы | |
| Символ | Pkinase_Tyr |
| Pfam | |
| Доступные структуры белков | |
| Pfam | |
| PDB | ; ; |
Рецепторные тирозинкиназы , рецепторы с тирозинкиназной активностью (англ. Receptor Tyrosin Kinases , сокр. RTK) — метаботропные клеточные рецепторы , представляют собой трансмембранные белки , которые состоят из каталитического внутриклеточного домена, участвующего в фосфорилировании субстратов ( тирозинкиназа ), трансмембранного и внешнего домена (связывающего) с высоким сродством к лигандам: гормонам ( инсулин ), цитокинам и полипептидным факторам роста . Из 90 уникальных генов тирозинкиназы, идентифицированных в геноме человека, 58 кодируют рецепторные тирозинкиназы .
Было показано, что рецепторные тирозинкиназы не только являются ключевыми регуляторами нормальных клеточных процессов, но также играют критическую роль в развитии и прогрессировании многих типов злокачественных опухолей . Мутации в рецепторных тирозинкиназах приводят к активации ряда сигнальных каскадов, которые оказывают многочисленные воздействия на экспрессию белка. Рецепторные тирозинкиназы являются частью более широкого семейства протеинтирозинкиназ, охватывающих рецепторные тирозинкиназные белки, которые содержат трансмембранный домен, а также нерецепторные тирозинкиназы, которые не обладают трансмембранными доменами .
Первыми RTK, которые обнаружили, были EGF (сокр. от англ. epidermal growth factor ) и NGF (сокр. от англ. nerve growth factor ) в 1960-х годах, однако классификация рецепторных тирозинкиназ не была разработана до 1970-х годов .
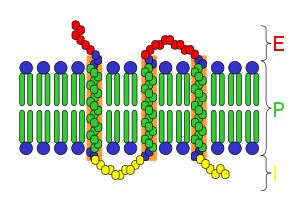
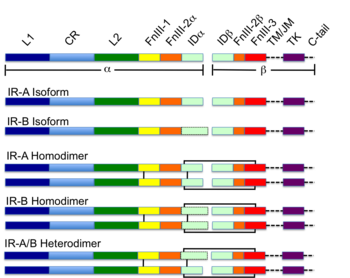
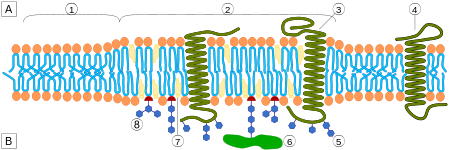
Большинство RTK являются рецепторами, состоящих из одной субъединицы, но некоторые существуют в виде мультимерных комплексов, например, инсулиновый рецептор , который образует дисульфид-связанные димеры в присутствии молекул гормона (инсулина); кроме того, связывание лиганда с внеклеточным доменом вызывает образование димеров рецептора . Каждый мономер имеет один гидрофобный трансмембранно-охватывающий домен, состоящий из 25-38 аминокислотных остатков, внеклеточную N-концевую область и внутриклеточную С-концевую область . Внеклеточная N-концевая область обладает множеством консервативных элементов, включая иммуноглобулин (Ig)-подобный или эпидермальный фактор роста (EGF)-подобные домены, повторы фибронектина типа III или богатые цистеином области, которые характерны для каждого подсемейства RTK; эти домены содержат в основном лиганд-связывающий сайт, который ответственен за связывание внеклеточных лигандов, например, факторов роста или гормонов . Внутриклеточная С-концевая область обладает наивысшим уровнем консервативности и содержит каталитические домены, ответственные за киназную активность этих рецепторов, которая катализирует аутофосфорилирование рецепторов и фосфорилирование остатков тирозина, являющихся субстратами RTK .
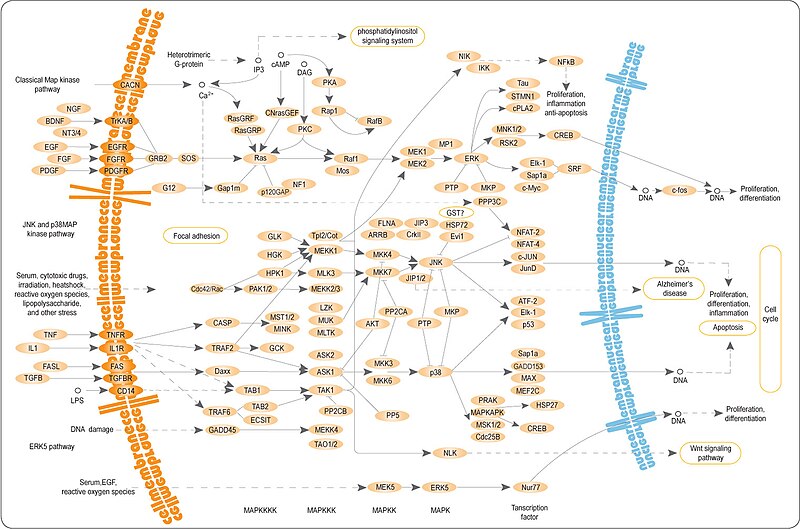
С помощью различных средств внеклеточное связывание лиганда обычно вызывает или стабилизирует димеризацию рецептора. Это позволяет трансфосфорилировать тирозин в цитоплазматической части каждого мономера рецептора его партнёрским рецептором, распространяя сигнал через плазматическую мембрану . Фосфорилирование специфических остатков тирозина в активированном рецепторе создает сайты связывания для белков, src-гомологичные домены 2 (SH2) и связывающий домен фосфотирозина (PTB) . Специфические белки, содержащие данные домены, включают Src и фосфолипазу Cγ. Фосфорилирование и активация этих двух белков при связывании рецептора приводят к инициации путей передачи сигнала. Другие белки, которые взаимодействуют с активированным рецептором, как адаптерные белки не имеют собственной ферментативной активности. Эти адаптерные белки связывают процесс активации молекул RTK с нисходящими сигнальными путями трансдукции, такими как сигнальный каскад MAP-киназы . Примером жизненно важного пути трансдукции сигнала является рецепторная тирозинкиназа, c-met, которая необходима для выживания и пролиферации мигрирующих миобластов во время миогенеза (роста и развития мышечной ткани). Недостаток c-met нарушает вторичный миогенез и как в LBX1-препятствует формированию мускулатуры конечностей. Такое локальное действие FGF (факторов роста фибробластов) с их рецепторами RTK классифицируется как паракринная передача сигналов. Поскольку молекулы RTK фосфорилируют множество остатков тирозина, они могут активировать множество путей передачи сигнала.
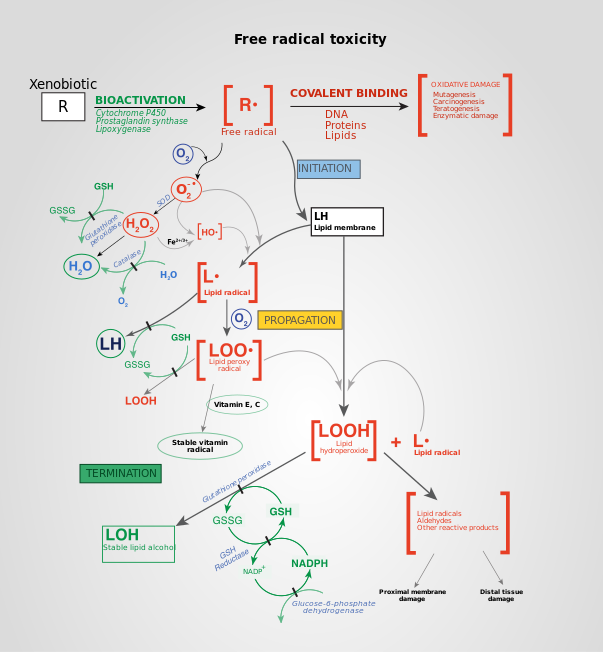
Рецепторный тирозинкиназный путь ( RTK pathway ) тщательно регулируется различными петлями положительной и отрицательной обратной связи . Поскольку RTKs координируют широкий спектр клеточных функций, таких как пролиферация и дифференцировка клеток , они должны регулироваться для предотвращения серьёзных нарушений в функционировании клеток, таких как рак и фиброз .
Белковые тирозинфосфатазы (англ. Protein Tyrosine Phosphatase , сокр. PTP) — это группа ферментов, обладающих каталитическим доменом с фосфотирозин-специфической фосфогидролазной активностью. PTP способны изменять активность рецепторных тирозинкиназ как в положительную, так и в отрицательную сторону . PTP могут дефосфорилировать активированные фосфорилированные остатки тирозина на RTKs , что практически приводит к прекращению передачи сигнала. Исследования с участием PTP1B, широко известного PTP, участвующего в регуляции клеточного цикла и сигнализации цитокиновых рецепторов, показали, что он дефосфорилирует рецептор эпидермального фактора роста и рецептор инсулина . Некоторые PTP, напротив, являются рецепторами клеточной поверхности, которые играют положительную роль в пролиферации клеточных сигналов. Cd45, гликопротеин клеточной поверхности, играет критическую роль в антиген-стимулированном дефосфорилировании специфических фосфотирозинов, ингибирующих путь Src .
Герстатин — аутоингибитор семейства рецепторов эпидермального фактора роста ErbB , связывающийся с RTK и блокирующий димеризацию и фосфорилирование рецепторов . В клетках CHO, трансфицированных герстатином, в ответ на EGF наблюдается снижение олигомеризации рецепторов, клонального роста и фосфорилирования RTK .
Активированные RTK могут подвергаться эндоцитозу , что приводит к снижению регуляции рецептора и, в конечном итоге, сигнального каскада . Молекулярный механизм включает в себя захват RTK клатрин-опосредованным эндоцитозом, что приводит к внутриклеточной деградации .
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)