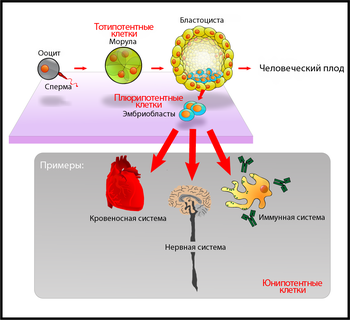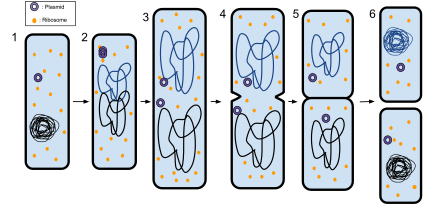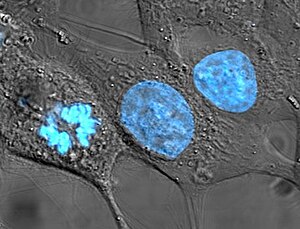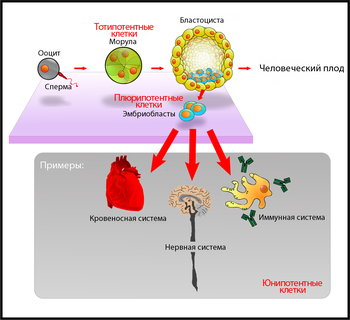
Дифференцировка клеток
- 1 year ago
- 0
- 0

Повреждение клеток (также известное как клеточное повреждение ) представляет собой различные изменения стресса, которым клетка подвергается из-за внешних, а также внутренних изменений окружающей среды. Среди других причин это может быть связано с физическими, химическими, инфекционными, биологическими, алиментарными или иммунологическими факторами. Повреждение клеток может быть обратимым или необратимым. В зависимости от степени повреждения клеточный ответ может быть адаптивным, и, по возможности, гомеостаз восстанавливается . Гибель клеток происходит, когда тяжесть повреждения превышает способность клетки к самовосстановлению . Гибель клеток зависит как от продолжительности воздействия вредного стимула, так и от тяжести причинённого ущерба . Гибель клеток может происходить путём некроза или апоптоза.
Наиболее заметными компонентами клетки, которые являются мишенями клеточного повреждения, являются ДНК и клеточная мембрана .
Некоторые повреждения клеток могут быть устранены после снятия стресса или при возникновении компенсаторных клеточных изменений. Полная функция может вернуться к клеткам, но в некоторых случаях степень повреждения останется .
Отек клеток (или тучный отек) может возникать из-за клеточной гипоксии , которая повреждает натрий-калиевый мембранный насос; оно обратимо при устранении причины . Отек клеток является первым проявлением почти всех форм повреждения клеток. Когда он поражает многие клетки в органе, он вызывает некоторую бледность, повышение тургора и увеличение веса органа. При микроскопическом исследовании в цитоплазме можно увидеть мелкие прозрачные вакуоли; они представляют собой расширенные и оторванные сегменты эндоплазматического ретикулума . Этот тип несмертельного повреждения иногда называют гидропическими изменениями или вакуолярной дегенерацией . Гидропическая дегенерация — тяжёлая форма тучного отека. Возникает при гипокалиемии вследствие рвоты или диареи.
Ультраструктурные изменения при обратимом повреждении клеток включают:
Повреждённая клетка не может адекватно метаболизировать жир. Мелкие жировые вакуоли накапливаются и диспергируются в цитоплазме. Умеренные жировые изменения могут не влиять на функцию клеток; однако более серьёзные жировые изменения могут нарушать клеточную функцию. В печени увеличение гепатоцитов из-за жировых изменений может сдавливать соседние , что приводит к холестазу . В зависимости от причины и степени накопления липидов жировые изменения, как правило, обратимы. Жировые изменения также известны как жировая дистрофия, жировая метаморфоза или жировой стеатоз.
Некроз характеризуется набуханием цитоплазмы, необратимым повреждением плазматической мембраны и разрушением органелл, что приводит к гибели клеток . Стадии клеточного некроза включают пикноз ; слипание хромосом и сморщивание ядра клетки , кариорексис ; фрагментация ядра и распад хроматина на бесструктурные гранулы, кариолизис ; растворение клеточного ядра . Цитозольные компоненты, просачивающиеся через поврежденную плазматическую мембрану во внеклеточное пространство, могут вызвать воспалительную реакцию .
Существует шесть типов некроза:
Апоптоз — это запрограммированная гибель лишних или потенциально вредных клеток в организме. Это энергозависимый процесс, опосредованный протеолитическими ферментами, называемыми каспазами, которые запускают клеточную гибель посредством расщепления специфических белков в цитоплазме и ядре . Отмирающие клетки сжимаются и конденсируются в апоптотические тельца. Поверхность клетки изменяется таким образом, чтобы проявлять свойства, которые приводят к быстрому фагоцитозу макрофагами или соседними клетками . В отличие от некротической гибели клеток, соседние клетки не повреждаются апоптозом, поскольку цитозольные продукты безопасно выделяются мембранами до того, как они подвергнутся фагоцитозу . Апоптоз считается важным компонентом различных биопроцессов, включая клеточный цикл, гормонозависимую атрофию, правильное развитие и функционирование иммунной и эмбриональной систем, а также способствует химической гибели клеток, которая является генетически опосредованной . Есть некоторые свидетельства того, что некоторые симптомы «апоптоза», такие как активация эндонуклеазы, могут быть ложно индуцированы без вовлечения генетического каскада. Также становится ясно, что митоз и апоптоз каким-то образом переключаются или связаны, и что достигнутый баланс зависит от сигналов, полученных от соответствующих факторов роста или выживания. Проводятся исследования, направленные на выяснение и анализ механизма клеточного цикла и сигнальных путей, которые контролируют остановку клеточного цикла и апоптоз . У среднего взрослого человека от 50 до 70 миллиардов клеток умирают каждый день из-за апоптоза. Ингибирование апоптоза может привести к ряду раковых, аутоиммунных заболеваний, воспалительных заболеваний и вирусных инфекций. Гиперактивный апоптоз может привести к нейродегенеративным заболеваниям, гематологическим заболеваниям и повреждению тканей.
При повреждении клетки, организм пытается восстановить или заменить её, чтобы продолжить нормальное функционирование. Если клетка умирает, организм удалит ее и заменит другой функционирующей клеткой или заполнит пробел соединительной тканью, чтобы обеспечить структурную поддержку оставшимся клеткам. Девиз процесса восстановления — заполнить пробел, вызванный повреждёнными клетками, чтобы восстановить структурную непрерывность. Нормальные клетки пытаются регенерировать повреждённые клетки, но это не всегда происходит. Бесполое размножение — это то, что восстанавливает клетки
Регенерация клеток паренхимы или функциональных клеток организма. Организм может производить больше клеток для замены повреждённых клеток, сохраняя орган или ткань неповреждёнными и полностью функциональными.
Когда клетка не может быть регенерирована, организм заменяет её стромальной соединительной тканью для поддержания функции ткани/органа. — это клетки, которые поддерживают паренхиматозные клетки в любом органе. Фибробласты, иммунные клетки, перициты и воспалительные клетки являются наиболее распространёнными типами стромальных клеток .
Истощение АТФ является распространённым биологическим изменением, которое происходит при повреждении клеток. Это изменение может произойти независимо от провоцирующего агента повреждения клетки. Снижение внутриклеточного АТФ может иметь ряд функциональных и морфологических последствий при повреждении клеток. Эти эффекты включают в себя:
Повреждение ДНК (или повреждение РНК в случае некоторых вирусных геномов) представляется фундаментальной проблемой для жизни. Как отмечает Хейнс , субъединицы ДНК не наделены какой-либо особой квантово-механической стабильностью, и поэтому ДНК уязвима ко всем «химическим ужасам», которые могут постигнуть любую такую молекулу в тёплой водной среде. Эти химические ужасы представляют собой повреждения ДНК, которые включают различные типы модификации оснований ДНК, одно- и двухцепочечные разрывы и межцепочечные перекрёстные связи (см. Повреждения ДНК (естественные) . Повреждения ДНК отличаются от мутаций, хотя и то и другое является ошибками в ДНК. В то время как повреждения ДНК представляют собой аномальные химические и структурные изменения, мутации обычно вовлекают нормальные четыре основания в новые структуры. Мутации могут реплицироваться и, таким образом, наследоваться при репликации ДНК. Напротив, повреждения ДНК представляют собой изменённые структуры, которые сами по себе не могут быть воспроизведены.
Несколько различных процессов репарации могут удалить повреждения ДНК (см. репарация ДНК ). Однако те повреждения ДНК, которые остаются невосстановленными, могут иметь пагубные последствия. Повреждения ДНК могут блокировать репликацию или транскрипцию генов. Эти блокировки могут привести к гибели клеток. У многоклеточных организмов гибель клеток в ответ на повреждение ДНК может происходить в результате запрограммированного процесса — апоптоза . В качестве альтернативы, когда ДНК-полимераза реплицирует цепь матрицы, содержащую поврежденный участок, она может неточно обойти повреждение и, как следствие, ввести неправильное основание, что приведёт к мутации. Экспериментально частота мутаций существенно возрастает в клетках, дефектных в репарации несоответствия ДНК или в гомологичной рекомбинационной репарации (HRR) .
Как у прокариот, так и у эукариот геномы ДНК уязвимы для атаки реактивными химическими веществами, естественным образом образующимися во внутриклеточной среде, и агентами из внешних источников. Важным внутренним источником повреждения ДНК как у прокариот, так и у эукариот являются активные формы кислорода (АФК), образующиеся как побочные продукты нормального аэробного метаболизма. Для эукариот окислительные реакции являются основным источником повреждения ДНК (см. Повреждение ДНК (природное) и Sedelnikova et al. . ). У человека ежедневно происходит около 10 000 окислительных повреждений ДНК на клетку . У крыс, у которых скорость метаболизма выше, чем у людей, на клетку приходится около 100 000 окислительных повреждений ДНК в день. У аэробно растущих бактерий АФК, по-видимому, являются основным источником повреждения ДНК, на что указывает наблюдение, что 89% спонтанно возникающих мутаций с заменой оснований вызваны введением индуцированных АФК одноцепочечных повреждений с последующей подверженной ошибкам репликацией после этих повреждений. повреждения . Окислительные повреждения ДНК обычно затрагивают только одну из цепей ДНК в любом поврежденном месте, но около 1–2% повреждений затрагивают обе цепи . Двухцепочечные повреждения включают двухцепочечные разрывы (DSB) и межцепочечные сшивки. Для человека предполагаемое среднее количество эндогенных DSB ДНК на клетку, возникающее в каждом поколении клеток, составляет около 50 . Такой уровень образования ДЦР, вероятно, отражает естественный уровень повреждений, вызываемых, в значительной степени, АФК, продуцируемыми активным метаболизмом.
Пять основных путей используются для восстановления различных типов повреждений ДНК . Этими пятью путями являются эксцизионная репарация нуклеотидов, эксцизионная репарация оснований, репарация несоответствия, негомологичное соединение концов и гомологичная рекомбинационная репарация (HRR) (см. репарация ДНК ) и источник . Только HRR может точно восстанавливать двухцепочечные повреждения, такие как DSB. Путь HRR требует, чтобы вторая гомологичная хромосома была доступна, чтобы позволить восстановить информацию, потерянную первой хромосомой из-за двухцепочечного повреждения.
Повреждение ДНК, по-видимому, играет ключевую роль в старении млекопитающих, а адекватный уровень восстановления ДНК способствует долголетию (см. и источник). Кроме того, повышенный риск повреждения ДНК и/или снижение репарации ДНК вызывают повышенный риск развития рака (см. Рак , канцерогенез и новообразования и источник ). Кроме того, способность HRR точно и эффективно восстанавливать повреждения двухцепочечной ДНК, вероятно, сыграла ключевую роль в эволюции полового размножения (см. и источник ). У современных эукариот HRR во время мейоза обеспечивает основное преимущество в поддержании фертильности .