Изоферменты
- 1 year ago
- 0
- 0
Глюкокиназа ( Шифр КФ ) представляет собой фермент, который способствует фосфорилированию глюкозы до глюкозо-6-фосфата . Глюкокиназа содержится в клетках печени и поджелудочной железы человека и большинства других позвоночных . В каждом из этих органов он играет важную роль в регулировании углеводного обмена , действуя как датчик глюкозы, вызывая сдвиги в метаболизме или функциях клеток в ответ на повышение или понижение уровня глюкозы, например, после еды или при голодании . Мутации гена этого фермента могут вызывать необычные формы диабета или гипогликемии .
Глюкокиназа (ГК) представляет собой изофермент гексокиназы , гомологично связанный по крайней мере с тремя другими гексокиназами . Все гексокиназы могут опосредовать фосфорилирование глюкозы до глюкозо-6-фосфата (Г6Ф), что является первой стадией как синтеза гликогена , так и гликолиза . Однако глюкокиназа кодируется отдельным геном , и её отличительные кинетические свойства позволяют ей выполнять другой набор функций. Глюкокиназа имеет более низкое сродство к глюкозе, чем другие гексокиназы, и её активность локализована в нескольких типах клеток, в результате чего остальные три гексокиназы являются более важными факторами подготовки глюкозы для гликолиза и синтеза гликогена для большинства тканей и органов. Из-за этого пониженного сродства активность глюкокиназы в обычных физиологических условиях существенно варьируется в зависимости от концентрации глюкозы .
Альтернативные названия этого фермента: человеческая гексокиназа IV, гексокиназа D и АТФ:D-гексоза 6-фосфотрансфераза, ЕС 2.7.1.1 (ранее 2.7.1.2). Общее название глюкокиназа происходит от её относительной специфичности к глюкозе в физиологических условиях.
Некоторые биохимики утверждают, что следует отказаться от названия глюкокиназа как вводящего в заблуждение, поскольку этот фермент может фосфорилировать другие гексозы в правильных условиях, а у бактерий есть отдалённо родственные ферменты с более абсолютной специфичностью для глюкозы, которые лучше заслуживают названия и EC от 19 октября 2003 на Wayback Machine . Тем не менее, название глюкокиназа остаётся предпочтительным в контексте медицины и физиологии млекопитающих.
Другая глюкозокиназа млекопитающих, , была открыта в 2004 году Этот ген отличается и похож на ген примитивных организмов. Он зависит от АДФ, а не от АТФ (что предполагает возможность более эффективного функционирования при гипоксии ), а метаболическую роль и важность ещё предстоит выяснить.
Основным физиологическим субстратом глюкокиназы является глюкоза , а наиболее важным продуктом — глюкозо-6-фосфат . Другой необходимый субстрат, из которого получают фосфат, — это аденозинтрифосфат (АТФ), который при удалении фосфата превращается в аденозиндифосфат (АДФ).
Реакция, катализируемая глюкокиназой:
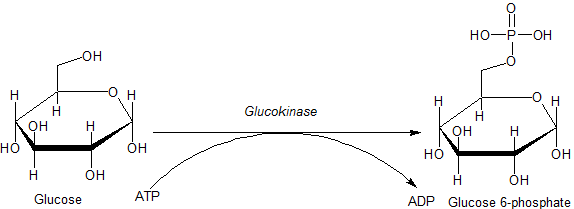
АТФ участвует в реакции в форме комплекса с магнием (Mg) в качестве кофактора . Кроме того, при определённых условиях глюкокиназа, как и другие гексокиназы, может индуцировать фосфорилирование других гексоз (6-углеродных сахаров ) и подобных молекул. Таким образом, общая реакция глюкокиназы более точно описывается как:
Среди субстратов гексозы есть манноза , фруктоза и глюкозамин , но сродство глюкокиназы к ним требует концентраций, не обнаруживаемых в клетках для значительной активности .
Два важных кинетических свойства отличают глюкокиназу от других гексокиназ, что позволяет ей выполнять особую роль в качестве сенсора глюкозы.
Эти две функции позволяют глюкокиназе регулировать метаболический путь, «управляемый поставками». То есть скорость реакции зависит от предложения глюкозы, а не от спроса на конечные продукты.
Ещё одним отличительным свойством глюкокиназы является её умеренная кооперативность с глюкозой с коэффициентом Хилла ( n H ) около 1,7 . Глюкокиназа имеет только один сайт связывания для глюкозы и является единственным мономерным регуляторным ферментом, который, как известно, проявляет кооперативность с субстратом. Постулируется, что природа кооперативности включает «медленный переход» между двумя различными состояниями фермента с разной скоростью активности. Если доминирующее состояние зависит от концентрации глюкозы, оно будет производить очевидную кооперативность, аналогичную наблюдаемой .
Из-за этой кооперативности кинетическое взаимодействие глюкокиназы с глюкозой не соответствует классической кинетике Михаэлиса-Ментен . Вместо K m для глюкозы более точно описать уровень половинного насыщения S 0,5 , который представляет собой концентрацию, при которой фермент является насыщенным и активным на 50 %.
S 0,5 и n H экстраполируются в «точку перегиба» кривой, описывающей активность фермента в зависимости от концентрации глюкозы около 4 ммоль/л. Другими словами, при концентрации глюкозы около 72 м/дл, что близко к нижнему пределу нормального диапазона, активность глюкокиназы наиболее чувствительна к небольшим изменениям концентрации глюкозы.
Кинетическая связь с другим субстратом, MgATP, может быть описана классической кинетикой Михаэлиса-Ментен со сродством примерно 0,3-0,4 ммоль/л, что значительно ниже типичной внутриклеточной концентрации 2,5 ммоль/л. Тот факт, что почти всегда имеется избыток доступного АТФ, означает, что концентрация АТФ редко влияет на активность глюкокиназы.
Максимальная удельная активность ( k cat , также известная как скорость оборота) глюкокиназы при насыщении обоими субстратами составляет 62 /с.
Оптимум pH глюкокиназы человека был идентифицирован только недавно, и он неожиданно высок и составляет 8,5-8,7 .
«Минимальная математическая модель» была разработана на основе вышеуказанной кинетической информации для прогнозирования скорости фосфорилирования глюкозы бета-клеток (BGPR) нормальной («дикого типа») глюкокиназы и её известных мутаций. BGPR для глюкокиназы дикого типа составляет около 28 % при концентрации глюкозы 5 ммоль/л, что указывает на то, что фермент работает на 28 % мощности при обычном пороговом уровне глюкозы для запуска высвобождения инсулина.
Сульфгидрильные группы нескольких цистеинов окружают сайт связывания глюкозы. Все, кроме Цис-230, необходимы для каталитического процесса, образуя множественные дисульфидные мостики во время взаимодействия с субстратами и регуляторами. По крайней мере, в бета-клетках соотношение активных и неактивных молекул глюкокиназы, по крайней мере, частично определяется балансом окисления сульфгидрильных групп или восстановления дисульфидных мостиков.
Эти сульфгидрильные группы весьма чувствительны к окислительному статусу клеток, что делает глюкокиназу одним из компонентов, наиболее уязвимых к окислительному стрессу, особенно в бета-клетках.
| Глюкокиназа | |
|---|---|
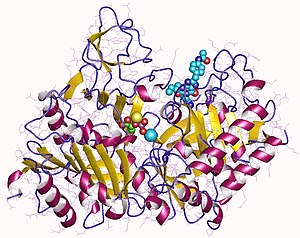
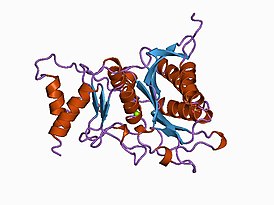
![Структура АТФ-зависимой глюкокиназы "Escherichia coli"[11].](/images/005/629/5629303/3.jpg?rand=856964)
Структура АТФ-зависимой глюкокиназы "Escherichia coli" . |
|
| Идентификаторы | |
| Pfam | |
| Pfam clan | |
| Доступные структуры белков | |
| Pfam | |
| PDB | ; ; |
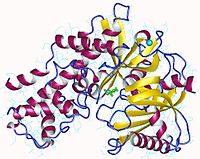
Глюкокиназа — это мономерный белок, состоящий из 465 аминокислот и молекулярной массы около 50 кДа . На поверхности есть по крайней мере два сайта, один для активного центра , в котором происходит связывание глюкозы и MgATP, а другой для предполагаемого аллостерического , который ещё не идентифицирован .
Это примерно вдвое меньше, чем у других гексокиназ млекопитающих, которые сохраняют определённую степень димерной структуры. Связывающий АТФ домен является общим с гексокиназами, бактериальными глюкокиназами и другими белками, и общая структура называется укладкой актина .
Глюкокиназа человека кодируется геном GCK на хромосоме 7 . Этот единственный аутосомный ген имеет 10 экзонов . Гены глюкокиназы у других животных гомологичны GCK человека .
Отличительной особенностью гена является то, что он начинается двумя промоторными участками . Первый экзон с 5'-конца содержит две тканеспецифичные промоторные области. Транскрипция может начинаться с любого промотора (в зависимости от ткани), так что один и тот же ген может продуцировать несколько разные молекулы в печени и в других тканях. Две изоформы глюкокиназы различаются только 13-15 аминокислотами на N-конце молекулы, что даёт лишь минимальное различие в структуре. Две изоформы имеют одинаковые кинетические и функциональные характеристики .
Первый промотор с 5'-конца, называемый «вышестоящим» или нейроэндокринным промотором, активен в клетках островков поджелудочной железы, нервной ткани и энтероцитах ( клетках тонкого кишечника ), продуцируя «нейроэндокринную изоформу» глюкокиназы . Второй промотор, «нижележащий» или печёночный промотор, активен в гепатоцитах и направляет продукцию «изоформы печени» . Два промотора имеют небольшую гомологию последовательностей или не имеют её и разделены последовательностью 30 тыс. пар оснований , которая, как ещё не было показано, вызывает какие-либо функциональные различия между изоформами . Два промотора функционально исключают друг друга и регулируются разными наборами регуляторных факторов, так что экспрессия глюкокиназы может регулироваться отдельно в разных типах тканей . Эти два промотора соответствуют двум широким категориям функции глюкокиназы: в печени глюкокиназа действует как ворота для «массовой обработки» доступной глюкозы, в то время как в нейроэндокринных клетках она действует как сенсор, запускающий клеточные реакции, которые влияют на организм: широкий углеводный обмен.
Глюкокиназа была обнаружена в определённых клетках четырёх типов тканей млекопитающих: печени , поджелудочной железы , тонкой кишки и мозга . Все они играют решающую роль в реагировании на повышение или понижение уровня глюкозы в крови .
Глюкокиназа печени встречается широко, но не повсеместно у позвоночных. Структура гена и аминокислотная последовательность высоко консервативны у большинства млекопитающих (например, глюкокиназа крысы и человека гомологична более чем на 80 %). Однако есть некоторые необычные исключения: например, она не была обнаружена у кошек и летучих мышей , хотя есть у некоторых рептилий , птиц , земноводных и рыб . Происходит ли аналогичное действие глюкокиназы в поджелудочной железе и других органах, ещё не установлено. Было высказано предположение, что присутствие глюкокиназы в печени отражает лёгкость, с которой углеводы могут быть включены в рацион животных.
Большая часть глюкокиназы у млекопитающих находится в печени, а глюкокиназа обеспечивает примерно 95 % активности гексокиназы в гепатоцитах. Фосфорилирование глюкозы до глюкозо-6-фосфата глюкокиназой является первым этапом как синтеза гликогена, так и гликолиза в печени.
Когда доступно достаточное количество глюкозы, синтез гликогена продолжается на периферии гепатоцитов до тех пор, пока клетки не наполнятся гликогеном. Затем избыток глюкозы всё больше превращается в триглицериды для экспорта и хранения в жировой ткани. Активность глюкокиназы в цитоплазме повышается и падает с доступной глюкозой.
Глюкозо-6-фосфат , продукт глюкокиназы, является основным субстратом синтеза гликогена, а глюкокиназа имеет тесную функциональную и регуляторную связь с синтезом гликогена. При максимальной активности глюкокиназа и по-видимому, расположены в тех же периферических областях цитоплазмы гепатоцитов, в которых происходит синтез гликогена. Поставка глюкозо-6-фосфата влияет на скорость синтеза гликогена не только в качестве основного субстрата, но и путём прямой стимуляции гликогенсинтазы и ингибирования гликогенфосфорилазы .
Активность глюкокиназы может быстро увеличиваться или подавляться в ответ на изменения в поставке глюкозы, обычно возникающие в результате приёма пищи и голодания. Регулирование происходит на нескольких уровнях и на нескольких скоростях, и на него влияют многие факторы, которые влияют в основном на два общих механизма:
Инсулин, действующий через белок −1c, связывающий регуляторный элемент стерола (SREBP1c), считается наиболее важным прямым активатором транскрипции гена глюкокиназы в гепатоцитах. SREBP1c — это (bHLHZ). Трансактиваторы этого класса связываются с последовательностью «Е-бокса» генов ряда регуляторных ферментов. Промотор печени в первом экзоне гена глюкокиназы включает такой E-бокс, который, по-видимому, является основным элементом инсулинового ответа гена в гепатоцитах. Ранее считалось, что SREBP1c должен присутствовать для транскрипции глюкокиназы в гепатоцитах, однако недавно было показано, что транскрипция глюкокиназы осуществляется нормально у мышей с нокаутом SREBP1c. SREBP1c увеличивается в ответ на высокоуглеводную диету, что, как предполагается, является прямым следствием частого повышения уровня инсулина. Повышенную транскрипцию можно обнаружить менее чем через час после воздействия на гепатоциты повышенного уровня инсулина.
Фруктозо-2,6-бисфосфат (Ф2,6БФ 2 ) также стимулирует транскрипцию ГК, по-видимому, посредством Akt2, а не SREBP1c. Неизвестно, является ли этот эффект одним из последующих эффектов активации рецепторов инсулина или не зависит от действия инсулина. Уровни F2,6P 2 играют другие усиливающие роли в гликолизе в гепатоцитах. 2 играют другие усиливающие роли в гликолизе в гепатоцитах. Другие факторы трансакции, которые, как предполагается, играют роль в регуляции транскрипции клеток печени, включают:
Инсулин , безусловно, является наиболее важным из гормонов, которые прямо или косвенно влияют на экспрессию и активность глюкокиназы в печени. Инсулин, по-видимому, влияет как на транскрипцию, так и на активность глюкокиназы множеством прямых и непрямых путей. В то время как повышение уровня глюкозы в воротной вене увеличивает активность глюкокиназы, сопутствующее повышение уровня инсулина усиливает этот эффект за счёт индукции синтеза глюкокиназы. Транскрипция глюкокиназы начинает расти в течение часа после повышения уровня инсулина. Транскрипция глюкокиназы становится практически невыявимой при длительном голодании, тяжёлой углеводной недостаточности или нелеченом инсулино-дефицитном диабете.
Механизмы, с помощью которых инсулин индуцирует глюкокиназу, могут включать как основные внутриклеточные пути действия инсулина, так и каскад киназ, регулируемых внеклеточными сигналами (ERK 1/2), и каскад фосфоинозитид-3-киназ (PI3-K). Последний может работать через трансактиватор FOXO1.
Однако, как и следовало ожидать, учитывая его антагонистический эффект на синтез гликогена, глюкагон и его внутриклеточный вторичный мессенджер цАМФ подавляют транскрипцию и активность глюкокиназы даже в присутствии инсулина.
Другие гормоны, такие как трийодтиронин (T 3 ), и глюкокортикоиды при определённых обстоятельствах оказывают разрешающее или стимулирующее действие на глюкокиназу. Биотин и ретиноевая кислота увеличивают транскрипцию мРНК GCK, а также активность GK. Жирные кислоты в значительных количествах усиливают активность GK в печени, в то время как ингибирует её.
Глюкокиназа может быть быстро активирована и инактивирована в гепатоцитах новым регуляторным белком (регуляторным белком глюкокиназы — ), который поддерживает неактивный резерв ГК, который может быстро стать доступным в ответ на повышение уровня глюкозы в воротной вене .
перемещается между ядром и цитоплазмой гепатоцитов и может быть привязан к цитоскелету микрофиламентов. Он образует обратимые комплексы 1:1 с ГК и может перемещать его из цитоплазмы в ядро. Он действует как конкурентный ингибитор глюкозы, так что активность фермента снижается почти до нуля при связывании ГК: комплексы ГКРБ секвестрируются в ядре, в то время как уровни глюкозы и фруктозы низкие. Секвестрация ядра может служить для защиты ГК от деградации цитоплазматическими протеазами . ГК может быстро высвобождаться из ГКРБ в ответ на повышение уровня глюкозы. В отличие от ГК в бета-клетках, ГК в гепатоцитах не связан с митохондриями.
Фруктоза в крошечных (микромолярных) количествах (после фосфорилирования до (Ф1Ф)) ускоряет высвобождение ГК из ГКРБ. Эта чувствительность к присутствию небольшого количества фруктозы позволяет ГКРБ, ГК и кетогексокиназе действовать как «система чувствительности к фруктозе», которая сигнализирует о том, что смешанная углеводная еда переваривается, и ускоряет утилизацию глюкозы. Однако фруктозо-6-фосфат (Ф6Ф) усиливает связывание ГК с помощью ГКРБ. Ф6Ф снижает фосфорилирование глюкозы ГК при гликогенолизе или глюконеогенезе .Ф1Ф и Ф6Ф связываются с одним и тем же сайтом на ГКРБ. Предполагается, что они продуцируют 2 разные конформации ГКРБ, одна способна связывать ГК, а другая нет.
Хотя большая часть глюкокиназы в организме находится в печени, меньшие количества в бета- и альфа-клетках поджелудочной железы, некоторых нейронах гипоталамуса и определённых клетках (энтероцитах) кишечника играют всё более важную роль в регуляции углеводного обмена. В контексте функции глюкокиназы эти типы клеток вместе называются нейроэндокринными тканями, и у них есть общие аспекты регуляции и функции глюкокиназы, особенно общий нейроэндокринный промотор. Из нейроэндокринных клеток бета-клетки островков поджелудочной железы являются наиболее изученными и изученными. Вероятно, что многие из регуляторных отношений, обнаруженных в бета-клетках, будут также существовать в других нейроэндокринных тканях с глюкокиназой.
В островковых бета-клетках активность глюкокиназы служит основным регулятором секреции инсулина в ответ на повышение уровня глюкозы в крови. По мере потребления Г6Ф увеличивающееся количество АТФ запускает серию процессов, которые приводят к высвобождению инсулина. Одним из непосредственных последствий учащённого клеточного дыхания является повышение концентраций НАДН и НАДФН (совместно именуемые НАД(Ф)Н). Этот сдвиг в окислительно-восстановительном статусе бета-клеток приводит к повышению уровня внутриклеточного кальция , закрытию каналов K- АТФ , деполяризации клеточной мембраны, слиянию секреторных гранул инсулина с мембраной и выбросу инсулина в кровь.
Именно в качестве сигнала для высвобождения инсулина глюкокиназа оказывает наибольшее влияние на уровень сахара в крови и общее направление метаболизма углеводов. Глюкоза, в свою очередь, влияет как на непосредственную активность, так и на количество глюкокиназы, продуцируемой бета-клетками.
Глюкоза немедленно усиливает активность глюкокиназы за счёт эффекта кооперативности.
Второй важный быстрый регулятор активности глюкокиназы в бета-клетках происходит за счёт прямого белок-белкового взаимодействия между глюкокиназой и «бифункциональным ферментом» ( фосфофруктокиназа-2 /фруктоза-2,6-бисфосфатаза), который также играет роль в регуляции гликолиза. Эта физическая ассоциация стабилизирует глюкокиназу в каталитически благоприятной конформации (несколько противоположной эффекту связывания ГКРБ), что усиливает её активность.
Всего за 15 минут глюкоза может стимулировать транскрипцию GCK и синтез глюкокиназы посредством инсулина. Инсулин вырабатывается бета-клетками, но часть его действует на рецепторы инсулина B-типа бета-клеток, обеспечивая усиление активности глюкокиназы с положительной обратной связью. Дальнейшая амплификация происходит под действием инсулина (через рецепторы А-типа) для стимуляции собственной транскрипции.
Транскрипция гена GCK инициируется через «вышестоящий», или нейроэндокринный, промотор. Этот промотор, в отличие от промотора печени, имеет элементы, гомологичные другим промоторам индуцированных инсулином генов. Среди возможных трансакционных факторов — Pdx-1 и PPARγ. Pdx-1 представляет собой фактор транскрипции гомеодомена, участвующий в дифференцировке поджелудочной железы. PPARγ — это ядерный рецептор, который реагирует на препараты глитазона повышением чувствительности к инсулину.
Бо́льшая часть, но не вся глюкокиназа, обнаруженная в цитоплазме бета-клеток, связана с секреторными гранулами инсулина и митохондриями. «Связанная» доля быстро падает в ответ на повышение секреции глюкозы и инсулина. Было высказано предположение, что связывание служит цели, аналогичной цели печёночного регуляторного белка глюкокиназы, — защите глюкокиназы от деградации, так что она быстро становится доступной по мере повышения уровня глюкозы. Эффект заключается в усилении ответа глюкокиназы на глюкозу быстрее, чем это может сделать транскрипция .
Также было высказано предположение, что глюкокиназа играет роль в чувствительности к глюкозе альфа-клетками поджелудочной железы, но доказательства менее последовательны, и некоторые исследователи не нашли доказательств активности глюкокиназы в этих клетках. Альфа-клетки встречаются в островках поджелудочной железы, смешанные с бета-клетками и другими клетками. В то время как бета-клетки реагируют на повышение уровня глюкозы секрецией инсулина, альфа-клетки отвечают снижением секреции глюкагона. Когда концентрация глюкозы в крови падает до гипогликемического уровня, альфа-клетки выделяют глюкагон. Глюкагон — это белковый гормон, который блокирует действие инсулина на гепатоциты, вызывая гликогенолиз, глюконеогенез и снижая активность глюкокиназы в гепатоцитах. Степень, в которой подавление глюкозы глюкагона является прямым эффектом глюкозы через глюкокиназу в альфа-клетках или косвенным эффектом, опосредованным инсулином или другими сигналами от бета-клеток, всё ещё не определена.
В то время как все нейроны используют глюкозу в качестве топлива, некоторые чувствительные к глюкозе нейроны изменяют свою скорость возбуждения в ответ на повышение или понижение уровня глюкозы. Эти чувствительные к глюкозе нейроны сконцентрированы в основном в и гипоталамуса , которые регулируют многие аспекты гомеостаза глюкозы (особенно реакцию на гипогликемию), использование топлива, насыщение и аппетит , а также поддержание веса. Эти нейроны наиболее чувствительны к изменениям глюкозы в диапазоне 0,5-3,5 ммоль/л уровень глюкозы.
Глюкокиназа была обнаружена в головном мозге в основном в тех же областях, которые содержат нейроны, чувствительные к глюкозе, включая оба ядра гипоталамуса. Ингибирование глюкокиназы устраняет реакцию вентромедиального ядра на приём пищи. Однако уровень глюкозы в мозге ниже, чем в плазме, обычно 0,5-3,5. ммоль/л. Хотя этот диапазон соответствует чувствительности нейронов, чувствительных к глюкозе, он ниже оптимальной чувствительности к перегибу для глюкокиназы. Предположение, основанное на косвенных доказательствах, состоит в том, что нейрональная глюкокиназа каким-то образом подвергается воздействию уровня глюкозы в плазме даже в нейронах.
Хотя было показано, что глюкокиназа присутствует в определённых клетках (энтероцитах) тонкого кишечника и желудка, её функция и регуляция не изучены. Было высказано предположение, что и здесь глюкокиназа служит сенсором глюкозы, позволяя этим клеткам обеспечивать один из самых ранних метаболических ответов на поступающие углеводы. Предполагается, что эти клетки участвуют в функциях
Поскольку инсулин является одним из, если не самым важным, регуляторов синтеза глюкокиназы, сахарный диабет всех типов снижает синтез и активность глюкокиназы по ряду механизмов. Активность глюкокиназы чувствительна к окислительному стрессу клеток, особенно бета-клеток.
Обнаружено около 200 мутаций гена глюкокиназы человека GCK , которые могут изменять эффективность связывания и фосфорилирования глюкозы, увеличивая или уменьшая чувствительность секреции инсулина бета-клетками в ответ на глюкозу и вызывая клинически значимую гипергликемию или гипогликемию .
Мутации GCK снижают функциональную эффективность молекулы глюкокиназы. Гетерозиготность по аллелям со сниженной активностью ферментов приводит к более высокому порогу высвобождения инсулина и стойкой лёгкой гипергликемии. Это состояние называется диабетом 2 типа у молодых людей в зрелом возрасте ( MODY2 ). В самом последнем обзоре мутации GCK , наблюдаемой у пациентов, говорится о 791 мутации, из которых 489, как полагают, вызывают диабет MODY и, следовательно, снижают функциональную эффективность молекулы глюкокиназы .
Гомозиготность по аллелям GCK с пониженной функцией может вызвать тяжёлый врождённый дефицит инсулина, приводящий к стойкому неонатальному диабету .
Было обнаружено, что некоторые мутации усиливают секрецию инсулина. Гетерозиготность для увеличения функциональных мутаций снижает пороговое значение глюкозы, которое запускает высвобождение инсулина. Это создаёт гипогликемию различной формы, включая преходящий или стойкий , или возникающую в более старшем возрасте. В самом последнем обзоре мутаций GCK , которые наблюдались у пациентов, утверждалось, что 17 мутаций GCK вызывают гиперинсулинемическую гипогликемию .
Гомозиготность мутаций по усилению функции не обнаружена.
Несколько фармацевтических компаний исследуют молекулы, активирующие глюкокиназу, в надежде, что она будет полезна при лечении диабета 1 и 2 типа .
Beautiful structural pictures illustrating the conformational changes and potential regulatory mechanisms
| Гликолиз |
|
||||||||
|---|---|---|---|---|---|---|---|---|---|
| Только глюконеогенез |
|
||||||||
| Регуляция | |||||||||