Клеточная стенка
- 1 year ago
- 0
- 0


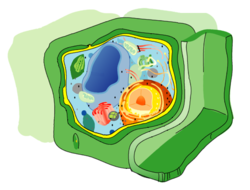
Кле́точная мембра́на (также цитолемма, плазмалемма, или плазматическая мембрана) — эластическая молекулярная структура, состоящая из белков и липидов . Отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы , в которых поддерживаются определённые условия среды.
Клеточная стенка , если таковая у клетки имеется (обычно есть у растительных, бактериальных и грибных клеток), покрывает клеточную мембрану.
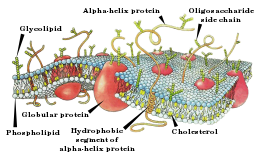
Клеточная мембрана представляет собой двойной слой ( бислой ) молекул класса липидов , большинство из которых представляет собой так называемые сложные липиды — фосфолипиды . Молекулы фосфолипидов имеют гидрофильную («головка») и гидрофобную («хвост») части. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь мембраны, а гидрофильные — наружу, с обеих внешних сторон мембраны. Важную роль в устойчивости внутренних участков мембран играют межмолекулярные взаимодействия , тогда как связность внешней и внутренней поверхностей мембран обеспечивается молекулами воды и субстрата, образующими с фосфатными группами фосфолипидов весьма прочные водородные связи в латеральной плоскости (вдоль поверхности мембраны). Мембраны — структуры , весьма сходные у разных организмов. Некоторое исключение составляют археи , у которых мембраны образованы глицерином и . Толщина мембраны составляет 7—8 нм .
Биологическая мембрана включает и различные белки : интегральные (пронизывающие мембрану насквозь), полуинтегральные (погружённые одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов , различных транспортеров и рецепторов .
В 1925 году и Франсуа Грендель (1897—1969) с помощью осмотического «удара» получили так называемые «тени» эритроцитов — их пустые оболочки. Тени сложили в стопку и определили площадь их поверхности. Затем с помощью ацетона выделили из оболочек липиды и определили количество липидов на единицу площади эритроцита — этого количества хватило на сплошной двойной слой. Хотя этот эксперимент привёл исследователей к правильному выводу, ими было допущено несколько грубых ошибок — во-первых, с помощью ацетона нельзя выделить абсолютно все липиды, а во-вторых, площадь поверхности была определена неправильно, по сухому весу. В данном случае минус на минус дал плюс, соотношение определяемых показателей случайно оказалось верным и был открыт липидный бислой.
Эксперименты с искусственными билипидными плёнками показали, что они обладают высоким поверхностным натяжением, гораздо большим, чем в клеточных мембранах. То есть в них содержится что-то, что снижает натяжение — белки. В 1935 году и представили научному сообществу модель «сэндвича», которая говорит о том, что в основе мембраны лежит липидный бислой, по обеим сторонам от которого находятся сплошные слои белков, внутри бислоя ничего нет. Первые электронно-микроскопические исследования 1950-х годов подтвердили эту теорию — на микрофотографиях были видны 2 электронно-плотных слоя — белковые молекулы и головки липидов и один электронно-прозрачный слой между ними — хвосты липидов. Дж. Робертсон сформулировал в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трёхслойное строение всех клеточных мембран.
Но постепенно накапливались аргументы против «бутербродной модели»:
Всё это привело к созданию в 1972 году и жидкостно-мозаичной модели строения мембраны. Согласно этой модели, белки в мембране не образуют сплошного слоя на поверхности, а делятся на интегральные , полуинтегральные и периферические. Периферические белки действительно находятся на поверхности мембраны и связаны с полярными головками мембранных липидов электростатичесткими взаимодействиями, но никогда не образуют сплошного слоя. Доказательствами жидкостности мембраны служат методы , и клеток, мозаичности — , при котором на сколе мембраны видны бугорки и ямки, так как белки не расщепляются, а целиком отходят в один из слоёв мембраны.

Мембраны состоят из липидов трёх классов: фосфолипиды , гликолипиды и холестерол . Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку.
Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются. Рядом с белками находятся аннулярные липиды — они более упорядочены, менее подвижны, имеют в составе более насыщенные жирные кислоты и выделяются из мембраны вместе с белком. Без аннулярных липидов белки мембраны не работают.
Клеточные мембраны часто асимметричны, то есть, слои отличаются по составу липидов, в наружном содержатся преимущественно фосфатидилинозитол , фосфатидилхолин , сфингомиелины и гликолипиды , во внутреннем — фосфатидилсерин , и фосфатидилинозитол . Переход отдельной молекулы из одного слоя в другой затруднён, но может происходить спонтанно, примерно раз в 6 месяцев или с помощью белков- флиппаз и плазматической мембраны. Если в наружном слое появляется фосфатидилсерин , это является сигналом для макрофагов о необходимости уничтожения клетки.
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы , отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть , аппарат Гольджи , лизосомы , вакуоли , пероксисомы ; к двумембранным — ядро , митохондрии , пластиды . Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза , аминокислоты , жирные кислоты , глицерол и ионы , причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия , осмос , активный транспорт и экзо - или эндоцитоз . Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия . При этом происходит дисбаланс мембранного потенциала, после чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия .
|
|
|
|---|---|
| Словари и энциклопедии |
|
| В библиографических каталогах |
|
Структуры
|
|
|---|---|
| Мембранные белки | |
| Другое | |
|
Эндомембранная система
|
|
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |
| Клеточная оболочка | |
|---|---|
| Наружная оболочка | |
| Форма | |
|
Термодинамические состояния
вещества
|
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Фазовые состояния |
|
||||||||||||||||
| Фазовые переходы |
|
||||||||||||||||
| Дисперсные системы | |||||||||||||||||
| См. также | |||||||||||||||||
Привет если ты видишь это знай у тебя всё получится!!!😁