Сумчатый волк
- 1 year ago
- 0
- 0

Берингийский волк — вымерший экоморф волка , обитавший на Земле в эпоху Валдайского оледенения . Населял современную Аляску , долину реки Юкон , северный Вайоминг . Часть волков данного подвида [ источник не указан 255 дней ] продолжала жить в голоцене .
В данный момент под вид [ источник не указан 255 дней ] всесторонне изучается рядом научных методов, дающих новую информацию. Выясняются сведения о рационе древних волков, их поведении. Было установлено, что данная группа морфологически отлична от современных волков Северной Америки и генетически она гораздо старше современных и вымерших волков. В данный момент берингийский волк не отнесён к какому-то подвиду, его классификация пока не уточнена. Связь с европейским пещерным волком ( Canis lupus spelaeus ) не выявлена .
Берингийский волк по размерам похож на современного юконского волка ( Canis lupus pambasileus ) и других серых волков, существовавших начиная с периода позднего плейстоцена. Его выделяют более сильные и прочные челюсти и зубы, более широкое нёбо и заметно более крупные, по сравнению с другими волками, хищные зубы , которые имеют размер больший, относительно размера черепа. Если сравнивать берингийского волка с ужасным волком , то ужасный волк (при одинаковом размере) имел более крепкий череп и зубы. Берингийский вид [ источник не указан 255 дней ] волка обладал уникальной адаптацией — сила его укуса позволяла охотиться на мегафауну ледникового периода, позволив ему занять нишу активного хищника, который вычищал плейстоценовую фауну от больных животных. Наиболее часто данный вид [ источник не указан 255 дней ] охотился на бизонов , диких лошадей , карибу , мамонтов и овцебыков .
С окончанием ледникового периода климат резко изменился, став более влажным, сократилось количество солнечных дней в году. Резко сократилась кормовая база для крупных травоядных. Потепление, исчезновение ледников, антропогенное воздействие времён палеолита и конкуренция с другими видами хищников привели к вымиранию берингийского волка. Его заместили волки одного вида и рода, но с другими генетическими особенностями, позволявшими им более эффективно охотиться и выживать в новом климате и при новых угрозах.
В популяции берингийских волков Северной Америки выжил только предок современного серого волка, который до сих пор обитает на континенте. Остатки древних волков со схожими черепами и зубной формулой были найдены в Западной Берингии (Северо-Восточная Сибирь). Исследования от 2015 года показали, что некоторые волки, живущие в наше время в отдалённых местах Китая и Монголии , имеют общего предка по материнской линии, который роднит их с берингийскими волками, обитавшими 28 000 лет назад .
В 2007 году Дженнифер Леонард провела анализ генетического материала из 74 образцов берингийского волка с Аляски и Юкона, который подтвердил, что это не один из типов серого волка, а по сути отдельный вид, имевший свои особенности в пищевом поведении, размерах черепа и нёба, строении зубов и генетике. Однако точного разделения на данном этапе исследований всё ещё не сделано. Поэтому отдельного, официального названия подвида берингийский волк ещё не получил. Это вызвано тем, что в прошлом Берингия соединяла широким перешейком Азию и Северную Америку, и животные могли свободно мигрировать между континентами. Отсюда схожесть в генетике у разных подвидов волков .
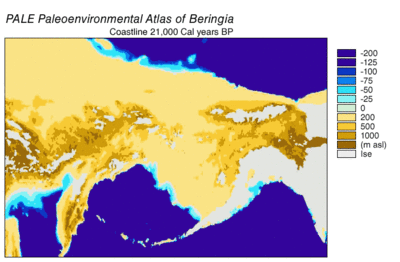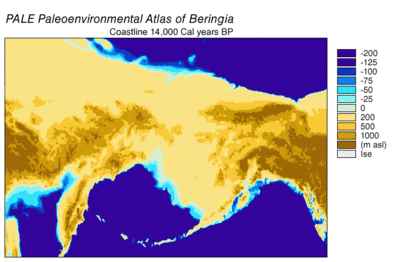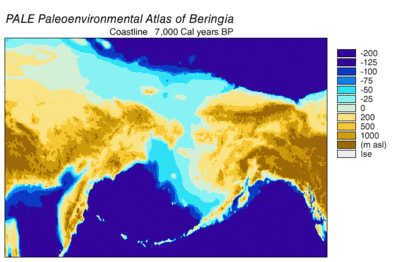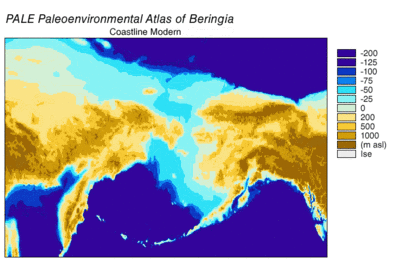
Секвенирование ДНК берингийского волка, показало, что он является генетическим предком для всех серых волков, живших после него, за исключением современного азиатского серого волка , гималайского волка и вымершей собаки времён палеолита, найденной в Бельгии . Генетическое разнообразие у подвидов берингийского волка было выше, чем у современных волков, в силу большей численности и большей занимаемой площади. Выяснилось, что современный волки на Аляске произошли не от берингийских волков, а от волков, мигрировавших из Евразии в период голоцена .
В соответствии с филогенетикой , размерами ледников и данными по уровню моря в разные эпохи была составлена следующая картина: во время позднего плейстоцена уровень моря был очень низким. Берингия соответственно увеличилась по площади своей суши, началась активная волна миграцией животных в Северную Америку. Первая такая волна произошла 70 000 лет назад. Затем, 23 000 лет назад, начался Последний ледниковый максимум , коридор вдоль долины реки Юкон оказался закрыт ледяным щитом ледника Лоурентида и Кордильерского ледника. Те животные, в том числе и волки, которые успели до оледенения проникнуть в Северную Америку, заменили собой имевшиеся виды волков, в том числе и за счёт своего большего количества. Ранняя популяция волков была полностью растворена .
| Phylogenetic tree for wolves | ||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||
| Филогенетическое древо Берингийских волков |

Берингийские волки, благодаря данным миграциям и замещению, морфологически и генетически сопоставимы с европейскими волками позднего плейстоцена . Так, при изучении находок волков в Одессе — 30 000 лет назад, в Крыму — 28 000 лет, в Алтайских горах 33 000 лет, в Чехии — 44 000 лет, выяснилось, что генетически по материнской линии они идентичны двум берингийским волкам. Берингийские волки связаны с отдельной группой из четырёх современных гаплотипов мтДНК (материнской) в Европе. Останки берингийских волков также идентичны находкам на Таймыре , на Украине , в Германии по строению черепов, зубов и челюстей. Учитывая, что в тот период тундростепь покрывала огромную площадь от нынешней Испании до Юкона на Аляске, мегафауна была наиболее вероятным источником пищи для волков и давала возможность мигрировать на столь большие расстояния. Приспособление волков к охоте на крупных животных привело в итоге к выделению отдельного вида [ источник не указан 716 дней ] — Canis cf. lupus , или Мегафаунный волк, специально адаптированный к охоте на мамонтов , зубров , овцебыков , гигантских оленей .
Случайные спаривания разных особей волков продолжались в Евразии и Северной Америке. Затем произошло перекрытие долины реки Юкон ледниками, после чего находившиеся южнее в Северной Америке волки оказались в изоляции. Берингийский волк оказался севернее образовавшегося ледяного щита и продолжал жить как в Северной Америке, так ив Евразии. 12 000 лет назад из-за поднятия уровня Мирового океана перешеек между континентами был затоплен. Ледяные щиты постепенно отступили между 14 000 — 8000 лет назад. Берингийский волк вымер, а популяция волков с Южной стороны Северной Америки стала расширятся и постепенно заселила площади, освобождённые ледником. Таким образом все современные североамериканские волки происходят от тех, которые остались южнее ледника .

По размерам берингийский волк совпадает с современным волком Юкона ( C. l. pambasileus ) — вес самцов достигает 43 кг, самок — 37 кг, средняя высота в холке — 80 см. Самые крупные современные волки на севере имеют высоту в холке до 97 см, длину тела — 180 см. В отдельных случаях встречаются мелкие особи — до 21 кг, и очень крупные — до 80 кг. Выделяется берингийский волк более прочными зубами и челюстью по сравнению с волками позднего плейстоцена и современными видами. Ужасный волк , обитавший южнее, имел среднюю массу 60 кг, достигая 68 кг. Он обладал ещё более крепким черепом и зубами .
Последний ледниковый период продолжался между 127—16,5 тыс. лет назад. Пик развития ледников и холодной погоды пришёлся на 35 тыс. лет назад и продолжался до 28,5 тыс. лет назад. Потепление и уменьшение площади ледников в Северном полушарии началось 21 тыс. лет назад, в Антарктиде — 16,5 тыс. лет назад. Подобная датировка согласуется с анализом фактов, указывающих на резкое повышение уровня Мирового океана 16,5 тыс. лет назад. В итоге 13 тыс. лет назад перешеек между Евразией и Северной Америкой оказался затоплен .
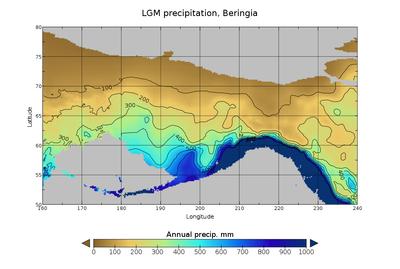
В Ледниковом периоде Тундростепь — занимавшая огромные площади от Испании до Юкона, от Арктических островов до Южного Китая, имела сухой и прохладный климат. 37000 лет назад Восточная Берингия имела температуру на 1,5 °C теплее, чем в современный период. 24 000 лет назад во время последнего ледникового максимума средняя температура была на 3—5 °C холоднее, чем современная. Берингия, в связи с близостью Тихого океана , имела больше облачных дней и больше осадков, чем остальная тундростепь. Подобные условия, связанные с большей влажностью, создавали зоны Рефугиума и позволили ряду растений и животных пережить похолодания климата. В изолированных карманах Берингии произрастали лиственничные и еловые леса с включениями берёзы и ольхи. Подобная среда позволял жить степным зубрам , мамонтам , юконским лошадям ( Equus lambei), якам — потреблявшим травы, осоки и травянистые растения; и северным оленям, и подвиду овцебыка — потреблявшим растения тундры, включая лишайники , грибы и мхи .
Анализ костей двух волков, найденных в Берингии и в Бельгии , позволил узнать рацион животных. Основа питания — животные мегафауны эпохи плейстоцена . Фауна начала сокращаться к началу голоцена , около 12 000 лет назад. Чаще всего волки охотились на семейство лошадиных и степных бизонов . Перед последним ледниковым максимумом 52 000 — 25 000 лет назад волки также охотились на овцебыков и мамонтов . Берингийские волки были способны убивать и расчленять очень крупных животных .
При анализе других костных останков половина волков питалась овцебыками и карибу . Вторая часть специализировалась на лошадиных и зубрах , однако могла охотится и на других зверей. По анализам останков двух волков обнаружено, что между 25—20 тыс. лет назад они охотились на мамонтов. Однако точно нельзя сказать, что стаи сами убивали животных. Возможно, они просто отбирали добычу у кого-то ещё. Мамонт , в силу размеров, был редкой добычей у волков как в Евразии , так и на Аляске .
По исследованиям 2007 года челюсти берингийских волков имели большие карнасальные зубы и короткое и широкое нёбо по сравнению с пропорциями черепа. Подобное соотношение делало череп и челюсти с зубами прочнее и давало возможность увеличивать силу укуса, оберегая кости от травм. Современные животные, обладающие схожими пропорциями и силой укуса, — пятнистые гиены . Гиены так же умеют охотиться на крупных животных, и устройство их черепов позволяет избегать травм, если добыча активно сопротивляется. Берингийские волки были гиперплотоядными животными. Подобная ориентация была вызвана наличием большой кормовой базы, которую давала мегафауна .
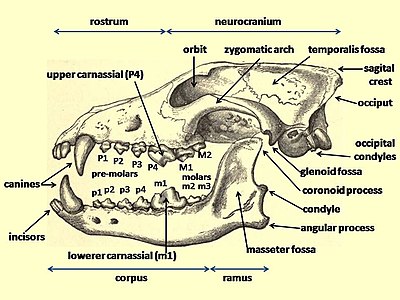
Согласно научной классификации, признаком одомашнивания является толщина зубов, их наклон, ориентация, плотность прилегания друг к другу .
| Обозначение зубов |
Совр. Сев.
Америка |
Ранчо
Ле Брей |
Берингия |
|---|---|---|---|
| Длина премолярной линии | 63,4 | 63,6 | 69,3 |
| Ширина нёба | 64,9 | 67,6 | 76,6 |
| Длина Р4 | 25,1 | 26,3 | 26,7 |
| Ширина Р4 | 10,1 | 10,6 | 11,4 |
| Длина М1 | 16,4 | 16,5 | 16,6 |
| Длина М2 | 8,7 | 8,9 | 9,2 |
| Длина m1 | 28,2 | 28,9 | 29,6 |
| Длина m1 моляров | 19,6 | 21,9 | 20,9 |
| Ширина m1 | 10,7 | 11,3 | 11,1 |

Исследования травм зубов вымерших волков и современного серого волка установили, что при разгрызании костей и отрывании кусков мяса от туши основную нагрузку несут моляры М2 и М3. Эти зубы используются в том числе для раскалывания костей. Современные серые волки обладают зубами с большей опорой в челюсти и, соответственно, большей крепостью и устойчивостью к повреждениям при разгрызании костей. У ужасного волка и других вымерших волков прочность крепления зубов была ниже. Серые волки в итоге лучше приспособлены для подобных действий и обладают меньшим процентом сломанных зубов. Современные северные волки Скалистых гор имеют в среднем 2% сломанных или повреждённых зубов, а берингийские волки имели 11% процентов повреждений. У берингийский волков значительно чаще обнаруживаются трещины и переломы резцов , хищных зубов и коренных зубов .
В период существования берингийских волков также жили пещерный лев , саблезубая кошка рода Homotherium , короткомордый медведь и всеядный бурый медведь . Активную конкуренцию волкам составлял именно короткомордый медведь , который питался падалью и активно искал убитых животных. Также 26 000 лет назад, люди достигли территории Юкона, об чём говорят останки степных зубров , карибу , вапити и барана Далла . Это повышало конкуренцию среди хищников, включая и человека. Животные активно поедали не только мясо, но и грызли кости ради костного мозга .
Тундростепь просуществовала благодаря периодическим оледенениям и сохранению большой площади ледников — 100 00 лет без особых изменений в климате и составе растительности. Закончился этот период 12 000 лет назад. В Северной Америке массовое вымирание животных весом больше 44 кг , то есть животных мегафауны, произошло начиная с 14 700 лет назад. Это связывают с активной охотой людей, которые, в силу улучшения климата, стали больше размножаться и проводить охотничью экспансию, и резким изменением климата, когда ледники стали таять, влажность повысилась и, следовательно, повысилось количество облачных дней в году — растения стали постепенно деградировать и тундростепь уступила место современной тундре и тайге . В общей сложности в Северной Америке вымерло 74% мегафауны . Под наибольшим ударом в вопросе питания оказались виды хищников, которые были полностью ориентированы на мясную пищу. В итоге берингийский волк, пещерный лев , саблезубые кошки рода смилодоны ( Smilodon ) и гомотерии ( Homotherium ), короткомордый медведь вымерли из-за узкой пищевой специализации и вымирания животных мегафауны — основы их питания .
Сумели выжить и дальше развиваться животные универсального типа питания — бурый медведь , американский чёрный медведь , койот , пума , рыжая рысь . Берингийский волк и ужасный волк вымерли в Северной Америке. Их сменил обладавший большей приспособленностью и универсальностью подвид серого волка . Согласно исследованиям 56 останков берингийских волков подтверждается непрерывная генетическая линия длительностью начиная с 52 800 лет назад и вплоть до 14 500 лет назад. После этого промежутка найден лишь один берингийский волк, датированный возрастом 9 600 лет назад. К этому времени вид почти полностью вымер. Сроки вымирания лошадей в Северной Америке и сокращения до минимум популяции бизонов коррелируют с вымиранием берингийских волков, точнее произошло прерывание единой генной линии вида, что свидетельствует о резкой убыли особей и невозможности поддерживать вид. В итоге волки унифицировались, значительно потеряв в своём геноме и фенотипическом многообразии по сравнению с периодом до вымирания берингийского и ужасного волков .